Lekcija 10 biologija 7. razred
T jesti O: Reprodukcija hidre. Regeneracija. Značenje u prirodi.
Zadatak.
Saznajte strukturne karakteristike i životne procese hidre kao niže višećelijske životinje.
Proučite karakteristike načina života u vezi sa staništem.
Razviti znanje o klasifikaciji hidra.
Formiranje vještina za rad sa mikropreparatima.
Oprema za nastavu.
Tabela „Slatkovodna Hidra, multimedijalni projektor, , mikroskopi, mikroslajd “Hydra”.
Ažuriranje znanja.
Navedite nivoe organizacije žive prirode. Kojem nivou pripadaju koelenterati i slatkovodna hidra? Kako se to može dokazati?
Koje vrste simetrije su karakteristične za životinje? Imenujte vrstu simetrije koelenterata.
Objasnite prednost ove vrste simetrije za koelenterate.
Ime karakteristične karakteristike tip Coelenterates.
Učenje novog gradiva
Uvodna riječ nastavnika.
Prije više od dva i po vijeka, jedan mladić je došao iz Švicarske u Holandiju. Upravo je završio fakultetsko obrazovanje iz nauke. Budući da mu je trebao novac, odlučio je da se zaposli kao tutor kod određenog grofa. Ovaj rad mu je ostavio vremena da sprovede vlastito istraživanje. Called mladi čovjek Abraham Tremblay. Njegovo ime je ubrzo postalo poznato širom prosvećene Evrope. I postao je poznat proučavajući ono što je doslovno svima bilo pod nogama - vrlo jednostavne organizme koji su živjeli u lokvama i jarcima. Tremblay je zamijenio jedno od ovih živih bića, koje je pažljivo pregledao u kapljicama vode izvučene iz jarka, za biljku.
Slajd 3.4.
Slatkovodna hidra pripada tipu coelenterate životinja. Među predstavnicima vrste koelenterata koji žive u morima, postoje sjedeći oblici - polipi i slobodno plivajući - meduze. Slatkovodna hidra je takođe polip.
Zapišite klasifikaciju vrste „Slatkovodna hidra“.
Aplikacija. Slajd 5
Vanjska struktura hidre
Hidrino tijelo u obliku tanke duguljaste vrećice, dugačke samo 2–3 mm do 1 cm, donjim je krajem pričvršćeno za biljku ili drugu podlogu. Donji dio tijelo se zove taban. Na drugom kraju hidrinog tijela nalaze se usta okružena vjenčićem od 6-8 pipaka.
Rad sa mikrouzorcima. Razmotrite vanjsku strukturu hidre.
Aplikacija. Slajd 6, 7
Nacrtajte vanjsku strukturu hidre u bilježnicu i označite dijelove tijela.
Ćelijska struktura hydra
Tijelo hidre ima oblik vrećice, čiji se zidovi sastoje od dva sloja ćelija: vanjskog - ektoderma i unutrašnjeg - endoderma. Između njih postoje slabi diferencirane ćelije. Šupljina koju formira ova vreća naziva se crijevna šupljina.
Aplikacija. Slajd 7, 8, 9.
Ispunjavanje dijagrama "Ektodermske ćelije"
Radimo samostalno. Popunite dijagram "Entodermalne ćelije"

Koji su vitalni procesi karakteristični za žive organizme?
Aplikacija. Hidra pokret. Slajd 13, 14.
Struktura nervnog sistema. Razdražljivost.
Aplikacija. Slajd 15,16.

Ishrana
Hidra je aktivan grabežljivac. Abram Tremblay je ovo rekao dok je posmatrao hidru.
Ako je hidra gladna, njeno tijelo se proteže cijelom dužinom, a pipci vise. Hrana koju hidra proguta iritira osjetljive ćelije endoderme. Kao odgovor na iritaciju, luče probavni sok u crijevnu šupljinu. Pod njegovim utjecajem dolazi do djelomične probave hrane.
Aplikacija. Slajd 17, 18.
Reprodukcija
Hidra se razmnožava spolno i aseksualno (pupanjem). Obično pupa u ljeto. Do jeseni se u tijelu hidre formiraju muške i ženske reproduktivne stanice i dolazi do oplodnje.

Aplikacija. Slajd 19, 20, 21.
Regeneracija
Abraham Tremblay je 25. septembra 1740. prepolovio hidru. Oba dijela su nastavili živjeti nakon operacije. Iz jednog komada, zvanog Tremblayeva "glava", izraslo je novo tijelo, a iz drugog - nova "glava". 14 dana nakon eksperimenta pojavila su se dva nova živa organizma. Hidra je mala, samo 2,5 centimetra. Tako malo stvorenje bilo je podijeljeno na stotinu dijelova - i iz svakog komada je nastala nova hidra. Podijelili su ga na pola i spriječili da polovice rastu zajedno - dobili su dvije životinje međusobno povezane. Hidra je secirana na snopove - formirana je kolonija hidra u obliku snopa. Kada je nekoliko hidri odrezano i pojedini dijelovi su ostavljeni da rastu zajedno, rezultat su bila potpuna čudovišta: organizmi s dvije glave, pa čak i nekoliko. I ovi monstruozni, ružni oblici su nastavili da žive, hrane se i razmnožavaju! Jedan od najpoznatijih Tremblayjevih eksperimenata je da je uz pomoć svinjske čekinje hidru okrenuo iznutra prema van, odnosno da je njena unutrašnja strana postala vanjska; nakon toga životinja je živjela kao da se ništa nije dogodilo.
Aplikacija. Slajd 22, 23, 24.

Konsolidacija.
Odaberite tačne izjave.
1. Među koelenterskim životinjama postoje predstavnici radijalne i bilateralne simetrije tijela.
Svi koelenterati imaju ubodne ćelije.
Svi koelenterati su slatkovodne životinje.
Vanjski sloj tijela koelenterata čine dermalno-mišićne, ubodne, živčane i srednje stanice.
Kretanje hidre nastaje zbog kontrakcije ubodnih niti.
Svi koelenterati su grabežljivci.
Koelenterati imaju dvije vrste probave - intracelularnu i ekstracelularnu.
Hidre nisu u stanju da reaguju na iritaciju.
2. Navedite karakteristične karakteristike slatkovodna hidra.
3. Popunite tabelu.
4. Upišite riječi koje nedostaju u rečenicama.Hidra je pričvršćena... za podlogu, na drugom kraju je..., okružena.... Hidra... organizam. Njegove ćelije su specijalizovane, formiraju... slojeve. Između njih je... . Posebnost koelenterata je prisustvo... ćelija. Naročito ih ima na... i oko usta. Spoljni sloj se zove..., unutrašnji sloj.... Kroz usta, hrana ulazi... u šupljinu.
Zadaća.
Proučite pasus.
Ponavljanje znakova koelenterata.
Pripremiti izvještaje o koelenterskim životinjama (meduze, koralji, morske anemone).
Hidrino tijelo izgleda kao duguljasta vreća, čiji se zidovi sastoje od dva sloja ćelija - ektoderm I endoderm.
Između njih leži tanak želatinozni nećelijski sloj - mesoglea, koji služi kao podrška.

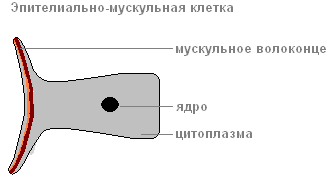
Ektoderm čini pokrivač životinjskog tijela i sastoji se od nekoliko vrsta ćelija: epitelno-mišićnog, srednji I peckanje.
Najbrojniji od njih su epitelno-mišićni.

Ektoderm
epitelne mišićne ćelije
Zahvaljujući mišićna vlakna, koje leži u dnu svake ćelije, tijelo hidre može se skupljati, produžiti i savijati.
Između epitelno-mišićnih ćelija nalaze se grupe malih, okruglih ćelija sa velikim jezgrima i malom količinom citoplazme, tzv. srednji.

Kada je hidrino tijelo oštećeno, oni počinju da rastu i brzo se dijele. Mogu se transformirati u druge vrste ćelija u tijelu hidre, osim u epitelno-mišićne.
Ektoderm sadrži ubodne ćelije, služi za napad i odbranu. Uglavnom se nalaze na pipcima hidre. Svaka ubodna ćelija sadrži ovalnu kapsulu u koju je umotana ubodna nit.
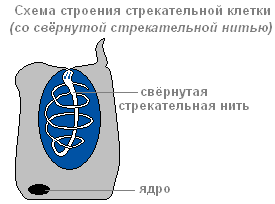
Struktura ubodne ćelije sa namotanom ubodnom niti
Ako plijen ili neprijatelj dotakne osjetljivu dlaku koja se nalazi izvan ćelije uboda, kao odgovor na iritaciju, ubodna nit se izbacuje i probija tijelo žrtve.
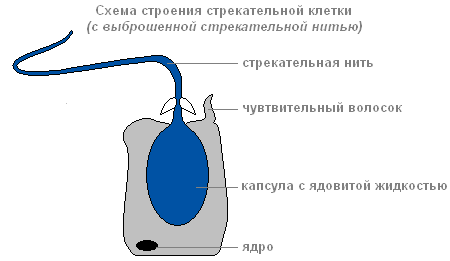
Struktura ubodne ćelije sa odbačenim ubodnim koncem
Kroz kanal navoja, supstanca koja može paralizirati žrtvu ulazi u tijelo žrtve.
Postoji nekoliko vrsta ubodnih ćelija. Konci nekog probijaju kožeživotinje i ubrizgavaju otrov u njihova tijela. Konci drugih su omotani oko plijena. Konci trećeg su veoma lepljivi i lepe se za žrtvu. Obično hidra "puca" nekoliko ubodnih ćelija. Nakon hica, ubodna ćelija umire. Od nje se formiraju nove ubodne ćelije srednji.
Struktura unutrašnjeg sloja ćelija
Endoderm oblaže cijelu crijevnu šupljinu iznutra. To uključuje digestivno-mišićni I žljezdanićelije.

Endoderm
Probavni sustav
Probavnih mišićnih ćelija ima više od ostalih. Mišićna vlakna sposobni su za redukciju. Kada se skrate, hidrino tijelo postaje tanje. Složeni pokreti (kretanje „tumbanjem”) nastaju zbog kontrakcija mišićnih vlakana ćelija ektoderma i endoderma.
Svaka od probavnih mišićnih ćelija endoderme ima 1-3 flagele. Oklevanje flagella stvaraju struju vode, koja tjera čestice hrane prema stanicama. Probavno-mišićne ćelije endoderme su sposobne da se formiraju pseudopodi, hvataju i vare male čestice hrane u probavnim vakuolama.
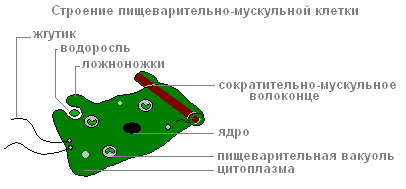
Struktura probavnih mišićnih ćelija
Imati u endodermu žlezdanih ćelija luče probavni sok u crijevnu šupljinu, koji ukapljuje i djelimično vari hranu.

Struktura ćelije žlezde
Plijen se hvata pipcima pomoću ubodnih ćelija, čiji otrov brzo paralizira male žrtve. Koordiniranim pokretima pipaka plijen se privodi ustima, a zatim se, uz pomoć tjelesnih kontrakcija, hidra „navlači“ na žrtvu. Varenje počinje u crijevnoj šupljini ( kavitetna probava), završava unutra digestivne vakuole epitelno-mišićne ćelije endoderme ( intracelularna probava). Hranljive materije se distribuiraju po celom hidrinom telu.
Kada se u probavnoj šupljini nalaze ostaci plijena koji se ne mogu probaviti i otpadni produkti staničnog metabolizma, ona se skuplja i prazni.
Dah
Hidra udiše kiseonik rastvoren u vodi. Ona nema respiratorne organe, a kiseonik upija po celoj površini tela.
Cirkulatorni sistem
Odsutan.
Odabir
Odabir ugljen-dioksid i drugih nepotrebnih materija koje nastaju u procesu života, prenosi se iz ćelija spoljašnjeg sloja direktno u vodu, a iz ćelija unutrašnjeg sloja u crevnu šupljinu, pa van.
Nervni sistem
Ispod ćelija kože i mišića nalaze se ćelije u obliku zvijezde. To su nervne ćelije (1). One se međusobno povezuju i formiraju živčanu mrežu (2).
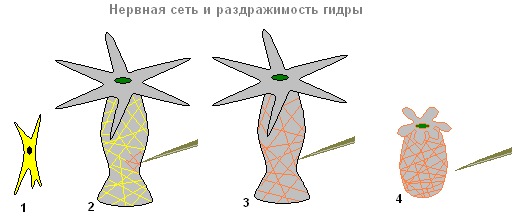
Nervni sistem i razdražljivost hidre
Ako dodirnete hidru (2), tada se u nervnim ćelijama javlja ekscitacija (električni impulsi), koja se trenutno širi po celoj nervnoj mreži (3) i izaziva kontrakciju kožno-mišićnih ćelija i skraćuje se celo telo hidre ( 4). Responsiveness hidrino tijelo na takvu iritaciju - bezuslovni refleks.
Polne ćelije
Sa približavanjem hladnog vremena u jesen, zametne ćelije se formiraju iz srednjih ćelija u ektodermu hidre.
Postoje dvije vrste zametnih stanica: jaja, ili ženske zametne stanice, i spermatozoidi, ili muške zametne stanice.
Jaja se nalaze bliže bazi hidre, spermatozoidi se razvijaju u tuberkulama koji se nalaze bliže ustima.
jajnu ćeliju Hidra je slična amebi. Opremljen je pseudopodima i brzo raste, upijajući susjedne međućelije.

Struktura jajne ćelije hidre

Struktura sperme hidre
Sperma po izgledu podsjećaju na bičaste protozoe. Napuštaju hidrino tijelo i plivaju koristeći dugačku bičicu.
Gnojidba. Reprodukcija
Spermatozoid sa jajnom ćelijom pliva do hidre i prodire u nju, a jezgra obje polne ćelije se spajaju. Nakon toga, pseudopodi se povlače, ćelija se zaokružuje, na njenoj površini se oslobađa debela ljuska - formira se jaje. Kada hidra umre i bude uništena, jaje ostaje živo i pada na dno. S početkom toplog vremena, živa ćelija koja se nalazi unutar zaštitne ljuske počinje se dijeliti, a rezultirajuće ćelije su raspoređene u dva sloja. Iz njih se razvija mala hidra, koja izlazi kroz puknuće ljuske jajeta. Dakle, višećelijska životinjska hidra na početku svog života sastoji se od samo jedne ćelije - jajeta. Ovo sugerira da su preci Hidre bili jednoćelijske životinje.

Aseksualno razmnožavanje hidre
At povoljnim uslovima Hidra se razmnožava aseksualno. Na tijelu životinje (obično u donjoj trećini tijela) formira se pupoljak, koji raste, zatim se formiraju pipci i probijaju se usta. Mlada hidra izbija iz majčinog tijela (u ovom slučaju polipi majke i kćeri se pipcima pričvršćuju za podlogu i uvlače u različite strane) i vodi samostalan način života. U jesen se hidra počinje razmnožavati spolno. Na tijelu, u ektodermu, nastaju spolne žlijezde - spolne žlijezde, a u njima se iz međućelija razvijaju zametne stanice. Kada se formiraju gonade hidre, formira se medusoidni čvor. Ovo sugerira da su gonade hidre vrlo pojednostavljeni sporifikatori, završna faza u seriji transformacije izgubljene medusoidne generacije u organ. Većina vrsta hidre je dvodomna; hermafroditizam je rjeđi. Jaja hidra rastu brzo fagocitozom okolnih ćelija. Zrela jaja dostižu prečnik od 0,5-1 mm. Oplodnja se događa u tijelu hidre: kroz posebnu rupu u gonadi, sperma prodire u jaje i spaja se s njim. Zigota je podvrgnuta potpunoj jednoličnoj fragmentaciji, zbog čega se formira coeloblastula. Zatim, kao rezultat mješovite delaminacije (kombinacija imigracije i delaminacije), dolazi do gastrulacije. Oko embrija formira se gusta zaštitna ljuska (embrioteka) sa izraslinama nalik na kičmu. U fazi gastrule, embriji ulaze u suspendovanu animaciju. Odrasle hidre umiru, a embrioni tonu na dno i prezimljuju. U proljeće se razvoj nastavlja, u parenhimu endoderme divergencijom stanica formira se crijevna šupljina, zatim se formiraju rudimenti pipaka, a ispod ljuske izlazi mlada hidra. Dakle, za razliku od većine morskih hidroida, hidra nema ličinke koje slobodno plivaju i njen razvoj je direktan.
Regeneracija
Hidra ima veoma visoku sposobnost regeneracije. Kada se poprečno preseče na više delova, svaki deo obnavlja „glavu“ i „nogu“, zadržavajući prvobitni polaritet – usta i pipci se razvijaju na strani koja je bila bliža oralnom kraju tela, a stabljika i taban se razvijaju na aboralna strana fragmenta. Cijeli organizam se može obnoviti iz pojedinačnih malih dijelova tijela (manje od 1/100 volumena), iz komadića pipaka, kao i iz suspenzije ćelija. Štoviše, sam proces regeneracije nije praćen povećanom diobom stanica i tipičan je primjer morfalaksije.
Pokret
U mirnom stanju, pipci se protežu nekoliko centimetara. Životinja ih polako pomiče s jedne strane na drugu, čekajući plijen. Ako je potrebno, hidra se može polako kretati.
"Pješački" način prijevoza

"Hodajući" način kretanja hidre
Zakrivivši svoje tijelo (1) i pričvrstivši svoje pipke na površinu predmeta (podloge), hidra povlači đon (2) na prednji kraj tijela. Zatim se ponavlja hodanje hidre (3,4).
"Tumbling" način kretanja

"Tumbling" metoda kretanja hidre
U drugom slučaju, čini se da se prevrće preko glave, naizmjenično se pričvršćujući pipcima i tabanom za predmete (1-5).
Kao što ste već shvatili iz prethodne prezentacije, mnogi predstavnici životinjskog carstva su sposobni za regeneraciju. Ali oblik i opseg regenerativnog rasta mogu se jako razlikovati među različitim životinjama. U ovom poglavlju upoznaćemo četiri poznata biologa kojima dugujemo mnogo našeg znanja o restauraciji izgubljenih organa. Svaki od ovih naučnika odabrao je svoj poseban put za proučavanje problema regeneracije i biće vam jasno da ne postoji jedan način da se problem reši. Razumijevanje mehanizma regeneracije može doći samo iz pažljivog poređenja informacija dobivenih korištenjem raznih eksperimentalnih pristupa.
ALLISON BURNETT. REGENERACIJA KOD HIDRE
Allison Burnett predaje na Univerzitetu Northwestern u Evanstonu, Illinois. Najveći dio svoje naučne aktivnosti posvetio je proučavanju ćelijske organizacije i procesa rasta v hydr. (Hydra), koji pripadaju istoj grupi beskičmenjaka kao i meduze, morske anemone i koralji. Kao što je Tremblay prvi primetio 1740. godine, regenerativna sposobnost hidra nije inferiorna po intenzitetu od regenerativne sposobnosti planarija. Stoga nije iznenađujuće da su studije regeneracije u hidrima i planarijama bile predmet stotina naučnih izvještaja na mnogim jezicima svijeta. Hidre su najčešći objekt za provođenje eksperimenata kako u obrazovne tako iu istraživačke svrhe.
Ove životinje slične biljkama obično žive u ribnjacima, pričvršćene za neku vodenu biljku ili stijenu pomoću ćelijskog diska (stopa) koji se nalazi u dnu tijela. Na suprotnom („glavnom“) kraju hidrinog tijela u obliku cijevi nalaze se usta koja se otvaraju u probavnu šupljinu nalik vrećici. Okružen je vjenčićem pipaka (od šest do deset), koji se neprestano kreću u potrazi za hranom. Jedan od načina na koji se hidre razmnožavaju je pupanjem - stvaranjem malih izbočina, ili pupoljaka, u donjem dijelu tijela beskičmenjaka. Postepeno, pipci i drugi organi tipični za hidre formiraju se na pupoljcima koji rastu. Tada se kćerka pojedinac odvaja od roditelja i započinje samostalan život. Nastali pupoljci daju hidri izgled višeglavog stvorenja. Visoka regenerativna sposobnost životinje poslužila je kao osnova da je dobila ime po starogrčkom mitološkom čudovištu, devetoglavoj Hidri, sposobnoj da lako obnovi glave odsječene u borbi (Sl. 33). Kod Hidra i srodnih životinja, tjelesni zid se sastoji od ćelija koje prekrivaju vanjski dio tijela (ektoderm) i imaju sposobnost kontrakcije, te stanica koje oblažu probavne šupljine (endoderma); prostor između ova dva sloja ispunjen je tankim slojem želatinozne supstance koja se zove mezoglea. Najčešće hidre ne prelaze 30 milimetara u dužinu.
E. Burnett je proučavao različita svojstva hidri. Svoje prve radove posvetio je proučavanju strukture i funkcije specijalizovanih ćelija ovih neobičnih životinja: nervnih ćelija koje stvaraju mrežu. nervne formacije u zidu tijela, karakteristično za sve koelenterate; ćelije žlezda koje čine endoderm i luče digestivni enzimi; kao i ubodne ćelije koje se nalaze na pipcima, sposobne da izbace uvrnutu nit s otrovom koji paralizira male životinje, plijen hidre, a u svrhu zaštite. Pored navedenih, male ćelije lišene ikakvih posebnih funkcija mogu se naći na mnogim mjestima u tijelu hidre; označene su kao intermedijarne, ili "intersticijske" ćelije ("I-ćelije"),
Hidra regeneracija
Burnett je svoje sljedeće eksperimente posvetio proučavanju posebnih faktora rasta za koje vjeruje da ih luči hidra. Postojanje takvih supstanci potkrijepljeno je opažanjima prirode regeneracije u hidri; Burnett i drugi istraživači su uspjeli otkriti da se zona rasta životinje nalazi u zidu tijela direktno ispod pipaka. Stalno stvaranje novih ćelija u ovoj zoni dovodi do toga da se obližnje zrele ćelije postepeno istiskuju u dva suprotna smera - prema pipcima i prema bazi tela - a nove ćelije, diferencirajući ih, zamenjuju ih. Kada "stare" ćelije stignu do krajeva hidrinog tijela, izbacuju se u okolno vodeno okruženje. Prema navedenoj hipotezi, ako neka od specijaliziranih ćelija umre prije završetka procesa migracije, njihovo mjesto zauzimaju obližnje I-ćelije, koje prolaze odgovarajuće promjene i preuzimaju funkcije zamijenjenih ćelija. Potonji način zamjene stanica se vrlo često opaža: ubodne ćelije se stalno troše u procesu hvatanja plijena, a žljezdane stanice - u procesu probave. Kao rezultat toga, hidrino tijelo prolazi gotovo kontinuirano obnavljanje (na oba načina), zbog čega je ova životinja, ne bez razloga, dobila ime "besmrtna".
Osim regenerativnih mehanizama koji rade kontinuirano, hidre se obnavljaju i kada su oštećene kao rezultat eksperimentalnih utjecaja. Ove životinje su sposobne ne samo regenerirati bilo koji od izgubljenih dijelova, već i potpuno obnoviti tijelo iz bilo kojeg najmanjeg fragmenta, s izuzetkom ticala i tabana. U procesu regenerativnog rasta primjećuje se jasan polaritet: kada se hidra vodoravno prepolovi, dio "glave", koji ima pipke, vraća stabljiku tabanom s površine rane, i obrnuto. Na prvi pogled, Hidra pokazuje gradijent svojstava duž linije ticala-stopalo, slično onome opisanom za planarijance. Burnett je, međutim, sugerirao drugačije. Nešto ranije, on i drugi istraživači došli su do zaključka da zona rasta ispod ticala luči posebnu tvar za rast, stimulativno proces ćelijska dioba. Sada je Burnett predložio da u istoj zoni, overwhelming rast je supstanca i da proces normalnog i regenerativnog rasta hidre zavisi od kombinacije ova dva faktora.
Hydra model rasta
Da bi ilustrirali valjanost svojih hipoteza, istraživači često pribjegavaju kreiranju modela određenih procesa. Burnettov model za regulaciju rasta hidre (slika 34) pretpostavlja da se tvari koje stimuliraju rast i inhibiraju rast polako kreću od mjesta proizvodnje prema dnu tijela životinje, a supstanca koja inhibira rast se sastoji od „tečnosti ” molekule koje postepeno izlaze iz tijela u okolinu.

Koje su karakteristike regeneracije hidre dale Burnettu osnovu za formulisanje principa rada njegovog modela? Prije svega, priroda regeneracije nakon disekcije. U gornjem dijelu, koji nosi pipke, proizvode se tvari koje potiskuju i stimuliraju rast. Logično je pretpostaviti da će doći do neutralizacije jednog faktora drugim. I doista, ne opažamo rast pipaka na raščlanjenom kraju, naprotiv, ovdje se počinje formirati stabljika s tabanom i vraća se polaritet karakterističan za tijelo životinje. Rast "glave" na površini rane donje polovice hidre potvrđuje još dva postulata hipoteze: prvo, u ovoj polovici hidre nema stanica koje bi mogle proizvesti tvar koja inhibira rast, i drugo , veći dio, koji je trebao doći do ovog dijela tijela, već je ispušten u okolinu.
Osim polariteta regenerativnog rasta Hydre, Burnettov model također objašnjava neke aspekte normalan oblik njegov rast, posebno reprodukcija pupoljkom. Sa stanovišta hipoteze o prisutnosti u tijelu Hidre gradijenta životnih procesa duž linije "pipci - taban", teško je razumjeti mehanizam pupanja. Prema gradijentnom modelu regeneracije u planarijama, brzina bilo kojeg biološkog procesa je mnogo veća na glavi životinje, a kod hidre brz rast, neophodan za pupanje, javlja se u dijelu tijela koji je veoma udaljen od “glave”. Ali, s druge strane, Burnettova teorija je ta koja lako objašnjava fenomen koji je uočen u prirodi. Važno je samo zapamtiti da je navodni faktor supresije rasta obdaren povećanom "fluidnošću". To stvara višak tvari za stimulaciju rasta u donjim dijelovima hidrinog tijela, što osigurava aktivan rast kćeri jedinki u području stabljike. U "bubrezima" ubrzo počinje samostalna proizvodnja tvari koja inhibira rast, što objašnjava polaritet tijela novoformiranih hidri.
Koje su najvažnije karakteristike Burnettovog modela regulacije rasta hidre? Objašnjava, prvo, normalne i regenerativne oblike rasta ovih beskičmenjaka uz pomoć jedne univerzalne teorije, i, drugo, uočeni polaritet rasta interakcijom dva specifična hemijska faktora. Ovo su izuzetno vrijedne ideje, ali ipak Burnettov model ne daje konačan odgovor na sva pitanja vezana za regeneraciju u Hidri. Njegov značaj je prvenstveno u tome što može poslužiti kao osnova za dalja eksperimentalna istraživanja koja trenutno provode kako sam Burnett, tako i drugi naučnici zainteresovani za ovaj problem.
MARCUS SINGER. ŽIVCI I REGENERACIJA
Već smo govorili o važnosti nerava u određenim fazama regeneracije udova kod vodozemaca. Marcus Singer sa Medicinskog fakulteta Univerziteta Cleveland, Ohajo, bio je prvi koji se zainteresovao za odnos između nervnog tkiva i regenerativnih procesa u različitim aspektima ovog problema.
U eksperimentima na denervaciji udova tritona, Singer je otkrio da regeneracija zavisi od očuvanja živca do faze formiranja dobro formiranog blastema panja. Niz daljnjih vrlo zanimljivih studija omogućio je Singeru da otkrije mogući način na koji nervno tkivo utiče na proces oporavka. Došao je do zaključka da nervno tkivo luči neku vrstu aktivna supstanca neophodna za regeneraciju. Singer govori o potrebi proučavanja ovog "neurotropnog" agensa na molekularnom nivou.
Kvalitet neophodnog nervnog tkiva
Svaki živac koji se nalazi u udovima kralježnjaka sastoji se od dva dijela. Jedan od njih - senzorni (osjetljivi) - prenosi nervne impulse od ekstremiteta do centralnog nervni sistem bez obzira na prirodu iritacije ekstremiteta. Drugi dio je motorni dio, prenosi signale iz centralnog nervnog sistema do mišića ekstremiteta, pružajući odgovor na različite vrste iritacija. Singer je prvo pokušao da utvrdi da li su oba dela živca uključena u obnavljanje udova tritona. Da bi to učinio, neposredno prije amputacije prednjeg ekstremiteta tritona, naučnik je presekao ili sve senzorne završetke tri glavna živca ekstremiteta, ili sve motorne (Sl. 35). Pokazalo se da regeneracija teče uspješno u obje varijante eksperimenta, odnosno kada je očuvana motorna ili senzorna inervacija. Iz ovoga možemo zaključiti da uticaj nervnog tkiva na regeneraciju nije visoka kvaliteta budući da vrsta nervnog vlakna koja ostaje u seciranom ekstremitetu ni na koji način ne utiče na njegovu regenerativnu sposobnost. Ali šta se može reći kvantitativno stranu stvari? Kako količina očuvanog nervnog tkiva utiče na proces regeneracije?

Potrebna količina nervnog tkiva
Analiza rezultata prethodnih eksperimenata pokazuje da za normalan oporavak udova nema potrebe za očuvanjem nervnog tkiva u uobičajenoj količini. Uostalom, potpuna obnova udova bez senzornih ili motornih nervnih završetaka događa se uz očigledan gubitak značajnog dijela živaca. Ali, pošto potpuno denervirani ekstremitet nije sposoban za regeneraciju, čini se da postoji određena minimalna količina nervnog tkiva neophodna za njegov regenerativni rast. Singer je predložio dizajn eksperimenata pomoću kojih je bilo moguće utvrditi vrijednost takvog minimuma.
I senzorni i motorički završeci tri glavna živca ekstremiteta sastoje se od snopova s određenim brojem nervnih vlakana međusobno povezanih vezivnim tkivom. U prvoj fazi eksperimenta određen je broj vlakana u svakom dijelu ova tri živca. Preparati poprečnih presjeka intaktnih nerava pripremljeni za mikroskopiju obojeni su na način da se može prebrojati broj vlakana u senzornoj i motornoj komponenti. Uz različite opcije za seciranje živaca kod eksperimentalnih životinja, lako je odrediti broj preostalih nervnih elemenata - da biste to učinili, samo trebate oduzeti broj seciranih od već poznatog broja vlakana određenog živca. Rezultati su bili prilično zanimljivi. Ako je u ekstremitetu ostalo više od 1298 nervnih vlakana, regeneracija je tekla normalno; ako je njihov broj pao ispod 793, do regeneracije nije došlo. Ako se broj očuvanih nervnih vlakana kretao od 793 do 1298, do restauracije ekstremiteta nekada je dolazilo, a nekada ne. Dakle, regeneracija se osigurava određenim prosječnim brojem nervnih vlakana (793-1298), tzv. nivo praga.
Logično je pretpostaviti da nedostatak sposobnosti regeneracije udova kod određene životinje može biti povezan s nedostizanjem graničnog broja nervnih vlakana. Ali daljnji eksperimenti Singera pokazali su da sposobnost regeneracije još uvijek nije određena ukupan broj nervna vlakna preostala nakon amputacije. Do ovog zaključka došao je upoređujući broj nervnih vlakana u udovima životinja brojnih vrsta. Kod životinja koje se ne mogu regenerirati, kao što su miševi ili odrasle žabe, dobiveni brojevi su bili znatno niži od graničnog broja za tritone. Ali brojeći broj nervnih vlakana Xenopus, Južnoafrička kandžasta žaba, neočekivano je pokazala da je jednako mali broj nervnih vlakana kod ovih životinja u kombinaciji sa dobro izraženom regenerativnom sposobnošću, koja se manifestuje i u odrasloj dobi (Sl. 36).

Ova kontradikcija je razriješena kada je, pored brojanja vlakana, određena veličina živaca udova kod predstavnika različitih grupa životinja. Ispostavilo se da nervnih vlakana at Xenopus značajno premašuju u prečniku ista vlakna kod miševa i odraslih žaba drugih vrsta. Kao rezultat toga, stepen inervacije ekstremiteta Xenopus znatno veći nego kod uporedivih životinjskih vrsta sa smanjenim regenerativnim kapacitetom. Pod uticajem dobijenih podataka bilo je potrebno malo promeniti koncept nivoa praga. Sada se kaže da udovi životinja imaju sposobnost regeneracije, pri čemu je osiguran određeni nivo snabdijevanja amputiranog područja ukupnom količinom nervnog tkiva, odnosno neuroplazma.
Prema Singeru, koncept nivoa praga uspješno objašnjava zašto se sposobnost obnavljanja udova smanjuje tokom evolucije, uprkos očiglednoj evolucijskoj "korisnosti" regeneracije. On tvrdi da kako je centralni nervni sistem postajao složeniji, došlo je do postepenog smanjenja količine nervnog tkiva u udovima. S tim u vezi, kod viših kičmenjaka nije postignut prag inervacije udova koji je pretpostavljen njegovom teorijom. Istovremeno, Singer smatra da nije uzalud priroda žrtvovala sposobnost obnavljanja udova, idući putem sve većeg poboljšanja centralnog nervnog sistema. Stjecanje sposobnosti donošenja brzih odluka koje omogućavaju životinji da se učinkovito brani od neprijatelja ima veću evolucijsku vrijednost od sposobnosti ponovnog rasta izgubljenih dijelova tijela.
Kako nervi stimulišu rast tkiva?
Sljedeća faza je bila proučavanje mehanizama utjecaja nervnog tkiva na regeneraciju. Singer je sugerisao da u ranim fazama regeneracije određena hemikalija koju luče nervi ima regulatorno dejstvo. Denervacija udova daždevnjaka tokom formiranja blasteme zaustavlja regeneraciju zbog činjenice da prestaje proizvodnja ove supstance. Šta ako, nakon denervacije, nekako nadoknadimo odsustvo navodnog hemijskog regulatornog faktora? Najteži dio ovih eksperimenata bio je pronaći način za liječenje denerviranih regenerirajućih udova daždevnjaka raznim kemikalijama. Pokušali smo nanijeti reagense direktno na površinu panja ili ih ubrizgati u tkivo pomoću šprice. Ali ni u jednom slučaju nije nastavljena regeneracija denerviranog panja. Takođe je bilo nemoguće isključiti mogućnost da hemijski stimulans sadržan u preparatima koji se koriste jednostavno nije dospeo do blastema, kao što se dešava kada ga u prirodnim uslovima luče nervi. Kako bi riješio ovaj problem, Singer je predložio poseban aparat, koji je trebao najpribližnije duplicirati normalnu aktivnost nerava, postupno otpuštajući ispitivane supstance direktno u blastemu ekstremiteta. Takav proces se zove infuzija, pa je stoga izum nazvan Singer aparat za mikroinfuziju.
Mikroinfuzija
Aparat koji je predložio Singer dizajniran je za kontinuirano protok malih količina tekućine kroz udove tritona, denerviranih u ranoj fazi regeneracije. Rad uređaja temelji se na rotaciji satnog mehanizma, koji se pretvara u translacijsko kretanje zavrtnja. Vijak, zauzvrat, pokreće mali klip hipodermički špric, isporukom otopine u tanku plastičnu cijev umetnutu umjesto igle. Slobodni kraj cijevi završava staklenom kapilarom, koja se nakon anesteziranja životinje ubacuje u područje ramena tritona, a zatim prodire u tkivo panja i blastema. Vijčani dio mehanizma može se povezati s pokretnom pločom koja pritiska od klipova brojnih špriceva - ova modifikacija omogućava simultanu infuziju na nekoliko tritona (Sl. 37).

Trigoni se drže pod anestezijom do pet sati i za to vrijeme razne hemikalije se unose u regenerirajući ekstremitet. Kako bi se osiguralo da se infuzija podudara prirodni proces oslobađanjem hemikalija nervnim tkivom, minimalne količine test rastvora se isporučuju u ekstremitet - oko 0,001 mililitara na sat.
Singer je sugerirao da najvjerovatnija supstanca koja utiče na regenerativnu sposobnost može biti neurotransmiter (transmiter nervne napetosti) acetilholin. Za ovu pretpostavku bilo je nekoliko razloga. Prvo, poznato je da acetilholin oslobađa nervno tkivo tokom prenosa impulsa. Drugo, pri određivanju sadržaja acetilholina u ekstremitetu u različitim fazama regeneracije, utvrđeno je da se u takozvanim živčano zavisnim fazama njegova količina pokazala većom u odnosu na normalno tkivo. Nakon formiranja blastema i tokom faze respecijalizacije, sadržaj acetilholina se vratio na normalan nivo(Sl. 38).

U regenerirajućim udovima tritona različiti periodi različite koncentracije acetilholina su davane tokom vremena. Činilo se vrlo vjerojatnim da bi infuzija acetilholina, barem u nekim slučajevima, pružila mogućnost regeneracije denerviranog ekstremiteta. Ali očekivanja nisu ispunjena. Infuzija denerviranih udova nikada nije završila proces regeneracije.
Unatoč izvjesnom razočaranju uzrokovanom rezultatom ovih eksperimenata, ipak ga treba smatrati vrijednim, jer eliminira jedan od mogući mehanizmi uticaj nervnog tkiva na regeneraciju i omogućava istraživačima da svoju pažnju usmere na potragu za alternativama.
Molekularna biologija i regeneracija ekstremiteta
Priroda supstance kojom nervno tkivo utiče na regeneraciju udova i dalje ostaje nejasna. Istraživači koji rade na ovom pitanju nedavno su pokušali da dešifruju mehanizam kojim nervi deluju na ćelije koje se regenerišu, nadajući se da će tako odrediti šta je uključeno u ovaj proces supstance.
Kada se ud denervira u fazi blastema, proces regeneracije se prekida, što ukazuje na prestanak aktivnosti ćelija blastema. Sasvim je logično pretpostaviti da denervacija utiče na jedan od najvećih važne funkcijećelije, odnosno proces sinteze proteina.
Detalji procesa sinteze proteina u ćelijama opisani su u bilo kom udžbeniku biologije, ali se mogu ukratko formulisati na sledeći način. Molekuli DNK, koji se nalaze u jezgru svake ćelije, sadrže kodirane informacije za sintezu različitih proteina. Oni služe kao svojevrsna matrica za formiranje molekula glasničke RNK, koji tu informaciju prenose do ribozoma smještenih u citoplazmi stanica. Tu se odvija proces sastavljanja proteina iz pojedinačnih "građevnih blokova", koji su aminokiseline. U eksperimentima koji su koristili radioaktivne izotope, Singer i njegove kolege pokušali su utvrditi kakav je učinak denervacija udova mrvica u ranoj fazi formiranja blastema imala na sintezu proteina u stanicama ekstremiteta. Pretpostavljali su da bi tokom denervacije ova vrsta ćelijske aktivnosti trebala prestati ili barem smanjiti.
Aminokiseline, kao i mnoge druge hemikalije, mogu se "označiti" ako se neki od elemenata zamijene radioaktivnim. Na osnovu intenziteta uključivanja označenih aminokiselina u proteinske molekule može se odrediti nivo sinteze proteina u ćelijama blastema. Shodno tome, efekat denervacije bi se trebao manifestirati u promjeni nivoa uključivanja označenih aminokiselina u proteine koje sintetiziraju ćelije blastema.
Dakle, označene aminokiseline su ubrizgane u tkivo regenerirajućih udova tritona. Za identifikaciju oznake u u ovom slučaju Koristili su metodu koja nije autoradiografija. U različito vrijeme nakon uvođenja označenih aminokiselina, dobijena je blastema, mljevena i izolirani proteini. Uzorci dobivenih materijala stavljeni su u scintilacijski brojač - uređaj koji vam omogućava da odredite razinu radioaktivnosti koju oslobađa određena količina proteina u minuti.
Rezultati eksperimenata potvrdili su početnu hipotezu: preparat proteina iz ćelija blastema denerviranih udova bio je značajno manje radioaktivan u odnosu na sličan preparat iz udova kod kojih je inervacija očuvana. Dakle, sinteza proteina u ćelijama blastema zaista zavisi od prisustva supstance koju luči nervno tkivo. Da bi potvrdio ovaj zaključak, Singer je sproveo eksperimente na kultivisanju nervnog tkiva i uneo materijal iz kultura u blasteme regenerisanih udova, koji su prethodno bili denervirani. Zatim je ponovio eksperiment sa radioizotopskom studijom sinteze proteina ćelijama blastema. Nakon infuzije materijala iz kultura nervnog tkiva, ćelije blastema denerviranih udova inkorporirale su obeležene aminokiseline sa približno istim intenzitetom kao ćelije blastema sa očuvanom inervacijom.
Dakle, prvi put je identifikovan na molekularnom nivou efekat supstance koju luči nervno tkivo koja utiče na proces regeneracije ekstremiteta. Ovo otkriće, značajno samo po sebi, ali i opšteg značaja, podstaklo je interesovanje naučnika za ovaj aspekt problema regeneracije i postavilo temelje za proučavanje specifičnih regulatornih mehanizama ovog biološkog fenomena.
ELIZABETH HAY. PROUČAVANJE PROCESA REGENERACIJE POMOĆU ELEKTRONSKOG MIKROSKOPA
Studije elektronske mikroskopije koje je provela Elizabeth Hay na Harvard Medical School mnogo su doprinijele našem razumijevanju sudbine pojedinačnih ćelija tokom regeneracije. Pomoću optičkog mikroskopa, kao što već znamo, utvrđen je opći slijed promjena na ćelijskom nivou koje se javljaju u regenerirajućim udovima daždevnjaka u fazama dediferencijacije, formiranja blastema i rediferencijacije. Međutim, ograničenje povećanja postignuto u optičkom mikroskopu ostavilo je mnoga pitanja bez odgovora. Na primjer, prilikom proučavanja regeneracije u planarijama, mikroskopija nije mogla odgovoriti da li do formiranja blastema dolazi zbog migracije rezervnih ćelija neoblasta ili zbog dediferencijacije zrelih ćelija. pljosnati crv. Pažljive elektronske mikroskopske studije koje je sproveo E. Hay omogućile su da se razriješi niz problema važna pitanja regeneraciju kod beskičmenjaka i kralježnjaka i dobiti puno dodatnih informacija.
Ćelije regenerirajućeg ekstremiteta
Koristeći konvencionalni optički mikroskop, ostala su neriješena tri glavna pitanja u proučavanju ćelija u regenerirajućim udovima daždevnjaka. Prvi od njih odnosio se na pojedinačne komponente ćelije, organele. Pretpostavljalo se da organele ćelija panja aktivno učestvuju u formiranju blastema. Ali kakvim promenama prolaze organele tokom dediferencijacije zrelih ćelija? Detalji ovog procesa ne mogu se proučavati optičkim mikroskopom.
Drugo pitanje je proizašlo iz podataka optičke mikroskopije, koji su pokazali da ćelije koje čine blastem ekstremiteta nemaju nikakve tragove porijekla iz jedne ili druge “roditeljske” ćelije i ne razlikuju se jedna od druge po strukturi. Da li su ćelije blastema zaista identične? Mnogi podaci su ukazivali na to, ali je ipak bilo nemoguće isključiti neke razlike koje su bile nevidljive u optičkom mikroskopu. Potrebne su dodatne studije kako bi se dokazalo da su ćelije blastema zaista izgubljene Sve strukturne karakteristike diferenciranih ćelija.
Treći problem se ticao nesigurnosti oko toga koja tkiva ekstremiteta podliježu dediferencijaciji i koja su dio blastema. Ovo se posebno odnosi na gubitak specijalizacije u mišićnom tkivu patrljka. Mikrofotografije dobijene optičkim mikroskopom pokazale su da se raščupani krajevi mišića ekstremiteta nakon amputacije u fazi „demontaže“, a neke mišićne ćelije u ovom području odvajaju od glavne mišićne mase, dediferenciraju i migriraju u Međutim, brojni istraživači su bili mišljenja da mišićno tkivo ne prolazi kroz proces dediferencijacije. Vjerovali su da nakon što se oštećeni krajevi intaktnih mišića oslobode ćelijskog detritusa, dolazi do direktnog ponovnog rasta novog mišićnog tkiva i prodiranja mišića u novoformirani dio ekstremiteta. Elektronska mikroskopska promatranja koje je proveo Hay omogućila su detaljnije proučavanje strukture citoplazme regenerirajućih stanica i dala odgovor na ova pitanja. Kao što ste vjerovatno već shvatili, u tu svrhu je korišten transmisioni elektronski mikroskop. Ultratanke presjeke normalnih i regenerirajućih udova aksolotla proučavane su s posebnom pažnjom na strukturu ćelija mišića i hrskavice, jer se te stanice u zrelom stanju lako prepoznaju po specifičnim supstancama koje luče.
Prije svega, utvrđena je priroda dvije vrste ćelija koje se spominju u neamputiranom udu. U citoplazmi zrelih ćelija hrskavice, pri velikim uvećanjima, jasno su se uočavale brojne membrane i ribozomi - male intracelularne čestice koje služe za sklapanje proteina iz aminokiselina. Ribosomi su bili u bliskoj vezi sa membranskim strukturama. Možete li se sjetiti drugog puta kada je sličan obrazac otkriven? Da, već smo vidjeli nešto slično na elektronskim mikrografijama fibroblasta uključenih u proces zacjeljivanja rana. Matrica koja okružuje ćelije hrskavice sadrži kolagen, kao i ožiljna tkiva formirana od fibroblasta, tako da oba tipa ćelija sintetiziraju molekule ovog proteina na ribozomima vezanim za membranu. U ćelijama hrskavice normalnog ekstremiteta nalazi se i Golgijev kompleks, tipičan za žljezdane ćelije. U zrelim mišićnim stanicama, gotovo cijeli prostor citoplazme zauzimaju snopovi kontraktilnog materijala, čija je poprečna pruga jasno vidljiva pod elektronskim mikroskopom.
Elektronske mikrografije su pokazale da se mišićne ćelije transformišu u tkivima blastema. U preparatima koje je Hay dobio u ranim fazama regeneracije, na mjestima disekcije mišićnog tkiva, među brojnim jezgrama preostalih intaktnih mišića, bile su vidljive granice novonastalih ćelija. Ovdje su pronađene i male ćelije, svaka od njih je imala po jedno jezgro. Kasnije su ove ćelije očigledno migrirale na površinu rane ekstremiteta i postale ćelije blastema.
Kada se pregledaju pod elektronskim mikroskopom, ćelije ranog blastema aksolotl ekstremiteta jasno su se razlikovale od zrelih ćelija mišića ili hrskavice (slika 39). Na primjer, citoplazmatske membrane ćelija blastema su bile fragmentirane, a ribosomi su bili slobodno razbacani po citoplazmi i nisu bili vezani za membrane. Iako je Golgijev kompleks u ćelijama blastema ostao uočljiv, bio je mnogo manji u poređenju sa Golgijevim kompleksom u zrelim ćelijama hrskavice. Citoplazma ćelija blastema bila je izuzetno slabo razvijena, ali jezgra su bila gigantske veličine i sadržavala su jasno definisane jezgre. Konačno, budući da proučavanjem ultrastrukture stanica blastema nisu otkriveni čak ni tragovi hrskavičnog matriksa ili mišićnih vlakana, zaključak dobiven optičkom mikroskopijom o identitetu stanica blastema je u potpunosti potvrđen.

Elektronske mikrofotografije snimljene tokom perioda rediferencijacije pokazale su da “pojednostavljene” organele ćelija blastema prolaze kroz postepene promjene kako se ud obnavlja, čija priroda je određena time koje specijalizirane ćelije nastaju na mjestu blastema. U centralno lociranim prekursorskim stanicama hrskavice postepeno se „pojavljuju“ citoplazmatske membrane sa vezanim ribosomima, Golgijev kompleks postaje sve izraženiji, a ubrzo se oko stanica počinje otkrivati ekstracelularni matriks. U prilično kasne faze regeneracije, kada su granice kostiju koje se oporavljaju već jasno vidljive, buduće mišićne ćelije koje se nalaze u vanjskim dijelovima blastema još ne pokazuju znakove respecijalizacije. Ali tada se pojavljuju ovi znakovi, stanice se produžuju, a kontraktilni materijal počinje se pojavljivati u citoplazmi. Čak i kasnije, ćelije se spajaju i formiraju tipičnu mišićno tkivo(Sl. 40). Tako je citološka studija faze respecijalizacije regenerirajućih udova aksolotla omogućila odgovor na sva tri pitanja postavljena na početku odjeljka.

Elektronska mikroskopija i regeneracija u planarijama
Mnogi istraživači su u optičkom mikroskopu otkrili grupe potpuno nespecijaliziranih ćelija smještenih u različitim dijelovima tijela ravnih crva. Ove ćelije nisu imale jasne razlike i razlikovale su se samo po prirodi bojenja njihove citoplazme određenim bojama. Budući da su migrirali prema površinama rane i učestvovali u formiranju blastema, nazvani su rezervnim ćelijama (neoblasti). Smatralo se da su neoblasti česti kod svih vrsta pljosnatih crva. Hay je nedavno proveo elektronsko mikroskopsko istraživanje ovih rezervnih ćelija u normalnim i regenerirajućim planarijama. Prva stvar koju je otkrila u ćelijama normalno ravnih crva, postoji značajan broj strukturnih detalja koji ukazuju na to da ćelije koje se proučavaju nisu bile nespecijalizovane u punom smislu te riječi. Velika povećanja elektronskog mikroskopa omogućila su da se u ovim ćelijama vide sekretorne granule i strukture Golgijevog kompleksa - jasne „granične stubove“ žlezdanih ćelija. Pojavila se pretpostavka da su rezervne ćelije namijenjene ne toliko da reagiraju na neke vrste oštećenja, već prije određenoj stalnoj funkciji - proizvodnji i izlučivanju sluzi. Sluz prekriva tijelo crva i omogućava mu da se kreće po raznim površinama koristeći mišićne kontrakcije.
U regenerirajuće Planarna elektronska mikroskopija otkrila je neobične ćelijske tokove usmjerene prema površini rane. Međutim, u tim tokovima nisu pronađene samo ćelije žlezda, već i niz drugih specijalizovanih ćelija. Kasnije, u blizini mjesta disekcije crva, migrirane stanice su postupno gubile svoje karakteristike specijalizacije, odnosno dediferencovale su se na potpuno isti način kao i stanice u regenerirajućim udovima vodozemaca. Po dolasku na površinu rane, sve migrirajuće ćelije su potpuno dediferencirane i spremne su za formiranje blastema. Neoblasti su stoga bili potpuno nepotrebni.
Kao što vidite, elektronska mikroskopija u mnogim slučajevima potvrđuje podatke zasnovane na opservacijama u optičkom mikroskopu. Ovo su bili rezultati detaljnog proučavanja ćelija regenerativnog ekstremiteta. Ali u primjeru planarnih rezervnih ćelija, elektronski mikroskopski podaci nisu se poklopili s rezultatima dobivenim ranije korištenjem manje napredne tehnologije. S tim u vezi, ponekad je potrebno preispitati objekte za koje se čini da su dugo proučavani, nakon čega se odredbe utvrđene u nauci često revidiraju.
RICHARD GOSS. RAZNOLIKOST SISTEMA REGENERACIJE
Richard Goss radi na Univerzitetu Brown u Providenceu, Rhode Island. Posvetio je svoj život proučavanju problema regeneracije raznih organa u veoma širokom spektru životinja. Ovdje ćemo predstaviti samo dva njegova rada, koja karakterišu izuzetnu širinu njegovih istraživačkih interesovanja. Govorit ćemo o regeneraciji antena okusa (mali brkovi osjetljivi izrasline koje okružuju usni otvor kod predstavnika nekih vrsta riba) i velikih razgranatih rogova kod jelena i losova, koji ponekad dosežu i 130 centimetara dužine.
Regeneracija okusnih pupoljaka
Som (na engleskom “catfish”, doslovno “mačka riba”) je dobio ime upravo po izuzetno izraženim antenama ukusa, koje veoma podsjećaju na mačji brkovi. Dr Goss je otkrio da kada se takva antena odseče od soma, na njenom mestu se formira blastema i izgubljeni proces se regeneriše. At mikroskopski pregled struktura antena se pokazala vrlo jednostavnom: svaka od njih sadržavala je živce i krvni sudovi, osnova organa bila je hrskavica, a na vrhu, ispod sloja epiderme, nalazio se okusni pupoljak.
Eksperimentalno istraživanje ovog minijaturnog sistema regeneracije otkrilo je niz zanimljivih činjenica. Blastema nastala nakon odsijecanja antena nastala je samo od dediferenciranih ćelija hrskavice. Ako je hrskavični štap uklonjen kroz mali rez na dnu antene, a zatim se sama antena presiječe, blastema se nije formirala i proces se nije regenerirao. Kako se pokazalo da je hrskavični štap neophodan za regeneraciju antene, logično je bilo pretpostaviti da ako se u jednu antenu stavi nekoliko šipki (moguće su i do četiri), onda nakon amputacije antene sa presjekom svih štapova, proces u nastajanju će sadržavati onoliko štapova koliko ih je bilo u panju. Ali eksperiment je otkrio samo jedan štap u regenerirajućoj vitici. Očigledno, gustatorna antenska blastema je "programirana" da formira normalan broj štapića u procesu, a prisustvo dodatnih struktura u panju ne utiče na normalan rast.
Regeneracija rogova jelena
Goss je potom svoju pažnju usmjerio na proučavanje regeneracije rogova kod jelena. Periodična prirodna zamjena ovih struktura je, po svemu sudeći, jedini primjer regeneracije tako složenog organa kod sisara. Ipak, pokazuje da je restauracija moguća i kod toplokrvnih životinja. velikih delova tijela. Stoga nije iznenađujuće što su mnogi istraživači pokazali veliko interesovanje za proučavanje ovog oblika regeneracije. Zbog ovoga opšti karakter Rast i sudbina pojedinih ćelija tokom restauracije rogova jelena, kao i činjenica o hormonskoj zavisnosti procesa regeneracije, prilično su dobro proučeni. Goss je, međutim, uspio da se otvori U poslednje vreme niz novih načina utjecanja na prirodne signale za stimulaciju hormonske aktivnosti tijela.
Tokom ranog perioda rasta mužjaka jelena, male koštane izrasline, ili panjevi, pojavljuju se na obje strane lubanje, nešto iznad i iza očiju. Kasnije se na tim mjestima formiraju mekani zaobljeni "napaljeni pupoljci", koji se potom izdužuju i granaju. Rast i razvoj roga odvija se od njegovog gornjeg kraja, ali okoštavanje hrskavičnih ćelija se odvija postepeno kako se udaljava od baze roga odozdo prema gore. Sličan gradijent diferencijacije tkiva uočen je sa svakim ciklusom regeneracije rogova.
Mužjaci jelena koji žive u umjerenom pojasu svake godine osipaju i ponovo uzgajaju svoje rogove, s prilično značajnim varijacijama kako u veličini rogova tako i u intenzitetu njihovog procesa regeneracije kod različitih vrsta. Goss je sastavio tabele koje pokazuju da veće životinje rastu brže. Elovi, najveći predstavnici porodice jelena, imaju rogove koji mogu doseći dužinu od 129,5 centimetara i rastu brzinom od 2,75 centimetara dnevno (Sl. 41, A, B). Kod svih jelena, kako rastući rog okoštava, krvne žile koje prodiru u njega bivaju začepljene koštanim tkivom, a koža s kratkom gustom dlakom ("smotulj") koja prekriva vanjsku stranu rogova, lišena opskrbe krvlju, puca i otpada. Otpadanje rogova, koji su se pretvorili u gustu kompaktnu koštanu masu, događa se mnogo kasnije, kada se na spoju rogova i panjeva pojavljuju ćelije, uništavajući koštane strukture. Rane brzo zacjeljuju i iznova počinje rast rogova. Kod većine vrsta opadanje rogova se dešava u kasnu zimu ili u proljeće, regeneracija se može vidjeti u ljetnim mjesecima, a opadanje sumota se događa neposredno prije sezone parenja, a to je u jesen. Svi ovi procesi su vođeni godišnjim fluktuacijama hormonske aktivnosti jelena. Smanjenje količine hormona testosterona u proljeće stimulira opadanje rogova i početak regeneracije, a povećanje njegovog nivoa u jesen dovodi do postepenog okoštavanja rogova i gubitka "smotulja".

Ako živite u srednjoj zoni, to znate različita vremena Dužina dnevnog svetla varira tokom godine. Ciklus rasta i promjene rogova hormonalni nivoi kod jelena je direktno povezan sa sezonskim promjenama dužine dana. Goss je u svojim eksperimentima stvorio režim umjetnog osvjetljenja kako bi dobio odgovore sledeća pitanja; prvo, da li je moguće promeniti ciklus zamene rogova produžavanjem ili skraćivanjem trajanja veštačkog svetla, i drugo, da li će do zamene rogova doći u uslovima kada je dužina dnevnog svetla konstantna tokom cele kalendarske godine?
U prvoj fazi svojih eksperimenata, izložio je jelene "svjetlosnoj godini" (kompletan godišnji ciklus povećanja i smanjenja dužine dana) koja je bila šest mjeseci van faze od uobičajene godine, što znači da su dani zimi postajali duži i kraći. ljeti. Pokusne životinje (sika jelen - mali, lagano građeni jeleni koji prirodno žive na Dalekom istoku, ali se nalaze u zoološkim vrtovima širom svijeta) držani su u negrijanoj prostoriji, gdje je temperatura bila podložna prirodnim godišnjim promjenama. Na rasvjetne uređaje bio je povezan poseban kalendarski mehanizam, uz pomoć kojeg se održavala „perverzija godišnjih doba“: u jesen, kako temperatura pada, dužina dnevnog vremena se postepeno povećava, dok se u proljeće, kako se temperatura zagrejana, dnevni sati skraćeni. Pri držanju u takvim uslovima, regeneracija rogova kod jelena jelena se dešavala u zimskim mesecima, a osipanje je bilo u jesen. Životinje su podvrgnute potpunoj adaptaciji na izopačene svjetlosne uvjete, barem što se rasta i regeneracije rogova tiče.
U nastojanju da u potpunosti testira svoju hipotezu, Goss je u narednim eksperimentima izložio životinje nekoliko ciklusa umjetne svjetlosti tokom jedne kalendarske godine. Da bi se to postiglo, kalendarski mehanizam je podešen tako da preskače svaki drugi dan. Time je osigurano da se izvode dva godišnja ciklusa promjene dnevnog vremena godišnje. Ako je mehanizam preskočio dva ili tri dana, godišnji ciklusi su se ponavljali tri ili četiri puta godišnje. U takvim uslovima jelen jelen je počeo da gubi rogove dva puta, tri puta ili četiri puta godišnje, prema broju eksperimentalnih godišnjih ciklusa. Zbog smanjenja ciklusa, dužina rastućih rogova je, naravno, bila znatno manja nego kod normalan ciklus(Sl. 42, A, B).

IN sledeći eksperiment Goss je produžio ciklus svjetlosne godine. Da bi se to postiglo, kalendarski mehanizam ponavljao se svakog normalnog dana dva puta, stvarajući tako „dvadesetčetvoromjesečnu godinu“. Sada je naučnik pokušavao da dobije odgovor na sledeća pitanja: da li će ciklus rasta rogova jelena koji se drže u takvim uslovima trajati sva 24 meseca i da li će to uticati na veličinu rogova ili ne? Odgovor na prvo pitanje pokazao se drugačijim u eksperimentima na jelenima različitog uzrasta: odrasle životinje nisu promijenile ciklus zamjene rogova, dok su se mladi jeleni lako prilagodili novoj dužini "godine", obnavljajući rogove samo jednom u gotovo dvije kalendarske godine. Što se tiče drugog pitanja, rast rogova koji prelazi normalnu dužinu nikada nije uočen, iako je ponekad bilo anomalija u prirodi njihovog rasta. Ali u svim slučajevima vještačkih promjena godišnjih doba, uočeni biološki efekti nisu bili trajni: većina jelena je po povratku u prirodne uvjete obnovila uobičajeni ciklični rast rogova.
Može li neki umjetno stvoren svjetlosni režim potpuno zaustaviti promjenu truba? U tu svrhu, eksperimentator je općenito isključio bilo kakve fluktuacije u dužini dnevnog svjetla. Niz godina je posebna grupa jelena držana u uslovima naizmjeničnog svjetla i mraka u tačno 12 sati. Takvi uslovi bili su identični onima koji su uočeni na ekvatoru. Svjetla su se palila u 6 sati ujutro i gasila u 6 sati uveče. U ovoj skupini većina jelena je potpuno izgubila sposobnost mijenjanja rogova, njihov ciklus regeneracije je potpuno poremećen. Dodatno, otkriveno je da je gubitak ciklusa povezan sa uporno povišenim nivoima testosterona.
Konačno, u završnom eksperimentu, Goss je izložio grupu jelena ciklusima nejednakih perioda svjetla i tame tokom kalendarske godine: osam, šesnaest ili dvadeset četiri sata svjetla praćenih šesnaest, osam sati tame ili bez svjetla. uopšte. U svakom takvom slučaju umjetno produženog ili skraćenog dnevnog svjetla, životinje su mogle s dovoljnom preciznošću odrediti stvarni protok vremena. Oni su mijenjali rogove jednom godišnje i iznenađujuće blizu vremena kada se ovaj proces odvija u prirodnim uslovima. Ovi rezultati snažno sugeriraju da jeleni imaju unutrašnji ritam kao što je "biološki sat". (U prethodnim Gossovim eksperimentima, takav ritam bio je podložan adaptivnim promjenama zbog umjetnih izopačenosti godišnjih ciklusa ili je bio potpuno poremećen kada su životinje prebačene u “ekvatorijalne” svjetlosne uslove, kada su se svjetlo i tama smjenjivali svakih 12 sati.) Šta god da je bilo. fiziološki mehanizam posmatran unutrašnji ritam, zavisi od glavnog faktora - nejednakosti trajanja svetlosnog i tamnog perioda u svakom 24-satnom ciklusu.
Iz rezultata koje su dobili drugi istraživači, čini se da tip odgovora koji je uočen kod jelena nije izuzetak. U veliki krug fiziološke promjene životinja i adaptivne reakcije usko su povezani sa promjenom dana i noći, promjenom godišnjih doba i izmjenom oseke i oseke. U mnogim slučajevima, kada su životinje uklonjene iz svog prirodnog staništa i lišene mnogih "znakova", one su ipak zadržale osjećaj za vrijeme i u skladu s tim zadržale svoje normalne biološke cikluse.
Rezultati Gossovih eksperimenata na regeneraciji rogova kod jelena sugeriraju da se regulacija drugih oblika može provesti na sličan način. procesi oporavka. Zaista, nedavno je naše znanje o fenomenima vezanim za funkcioniranje " biološki sat“, stalno se proširuju. I očigledno, nije daleko dan kada ćemo saznati kako se pokreće ovaj čudesni satni mehanizam prirode.
Biolozi koje smo sreli u ovom poglavlju nastavljaju svoja istraživanja o regeneraciji. Allison Burnett je u eksperimentima na hidrama razjasnila mehanizme regulacije rasta kod ovih koelenterata. Osim toga, neke od svojih teorija pokušava primijeniti na analizu regeneracije kičmenjaka. Budući da su osnovni obrasci aktivnosti stanica kod životinja različitih evolucijskih grupa izuzetno slični, prirodno je da stručnjaci za rast i razvoj jedne grupe životinja nastoje proširiti svoje zaključke na druge. Marcus Singer, utvrdivši da faktor koji oslobađa nervno tkivo može uticati na ćelijske mehanizme sinteze proteina na nekoliko načina, počeo je da traži biohemijsku „metu“ ove supstance u ćelijama blastema ekstremiteta. Među mnogim aspektima neurobiologije kojima se intenzivno bavi je i njegov Posebna pažnja privlači proučavanje strukture i funkcija mijelinske ovojnice aksona. Što se tiče Elizabeth Hay, njena vještina elektronskog mikroskopista sada služi ne samo zadacima proučavanja regeneracije. Ispitivala je finu strukturu širokog spektra embrionalnih ćelija - posebno ćelija srca i sočiva pilećeg embriona - i uporedila svoja zapažanja sa funkcijama ovih ćelija u procesu embrionalni razvoj. Richard Goss trenutno najviše pažnje posvećuje takvim procesima kompenzacijske regeneracije kao što je obnavljanje tkiva jetre i bubrega kod sisara nakon uklanjanja odgovarajućeg organa. Prema Gossu, povećanje znanja o ovim procesima će dovesti do otkrića specifičnih regulatora rasta tkiva i organa kod sisara.
Rad ovih naučnika – kako su upravo opisani, tako i koji se trenutno bave – predstavlja, naravno, samo dio te velike istraživačke aktivnosti koja je posvećena proučavanju regenerativnih procesa. Samo zajedno mogu dati holističku sliku svakog specifičnog procesa. Ali generalno gledano, izgledi su ohrabrujući. Regeneracija je postala važan dio razvojna biologija - grana nauke koja proučava obrasce normalnog i patološkog rasta, diferencijaciju ćelija, eksperimentalnu embriologiju i mnoge druge srodne probleme. Pogodne izglede za razvoj nove grane nauke određuje i činjenica da se posljednjih godina u nju kontinuirano slijevaju novi entuzijasti proučavanja regeneracije.
Jedan od tipičnih predstavnika reda koelenterata je slatkovodna hidra. Ova stvorenja žive u čistim vodenim tijelima i vezuju se za biljke ili tlo. Prvi ih je vidio holandski izumitelj mikroskopa i poznati prirodnjak A. Leeuwenhoek. Naučnik je čak uspio svjedočiti pupanju hidre i ispitati njene ćelije. Kasnije je Carl Linnaeus rodu dao naučno ime, pozivajući se na drevne grčke mitove o lernejskoj hidri.
Hidre žive u čistim vodenim tijelima i vežu se za biljke ili tlo.
Strukturne karakteristike
Ovaj vodeni stanovnik odlikuje se minijaturnom veličinom. U prosjeku, dužina tijela je od 1 mm do 2 cm, ali može biti i malo više. Stvorenje ima cilindrično tijelo. Ispred se nalaze usta sa pipcima (njihov broj može doseći i do dvanaest komada). Sa stražnje strane se nalazi đon, uz pomoć kojeg se životinja kreće i pričvršćuje za nešto.
Na tabanu se nalazi uska pora kroz koju prolaze mjehurići tekućine i plina iz crijevne šupljine. Zajedno sa mjehurićem, stvorenje se odvaja od odabranog nosača i lebdi. Istovremeno, glava mu se nalazi u gustoj vode. Hidra ima jednostavnu strukturu, tijelo joj se sastoji od dva sloja. Čudno, kada je stvorenje gladno, njegovo tijelo izgleda duže.
Hidre su jedni od rijetkih koelenterata koji žive u slatkoj vodi. Većina ovih stvorenja nastanjuje morsko područje . Slatkovodne vrste mogu imati sljedeća staništa:
- ribnjaci;
- jezera;
- riječne fabrike;
- rovovi.
Ako je voda bistra i čista, ova bića radije budu blizu obale, stvarajući neku vrstu tepiha. Drugi razlog zašto životinje preferiraju plitka područja je ljubav prema svjetlu. Slatkovodna stvorenja vrlo su dobra u razlikovanju smjera svjetlosti i približavanju njegovom izvoru. Ako ih stavite u akvarij, sigurno će doplivati do najosvijetljenijeg dijela.
Zanimljivo je da jednoćelijske alge (zoohlorela) mogu biti prisutne u endodermi ovog stvorenja. To se odražava na izgled životinje - dobiva svijetlozelenu boju.
Proces ishrane
Ovo minijaturno stvorenje je pravi grabežljivac. Vrlo je zanimljivo saznati čime se hrani slatkovodna hidra. Voda je dom mnogih malih životinja: kiklopa, cilijata i rakova. Oni služe kao hrana za ovo stvorenje. Ponekad može pojesti veći plijen, kao što su mali crvi ili larve komaraca. Osim toga, ovi koelenterati nanose veliku štetu ribnjacima, jer kavijar postaje jedna od stvari kojom se hidra hrani.
U akvariju možete u svom sjaju gledati kako ova životinja lovi. Hidra visi sa svojim pipcima nadole i istovremeno ih slaže u obliku mreže. Njen torzo se lagano njiše i opisuje krug. Plijen koji pliva u blizini dodiruje pipke i pokušava pobjeći, ali iznenada prestaje da se kreće. Ubodne ćelije paralizirati je. Zatim ga čoelenteratno stvorenje povuče ustima i pojede.
Ako je životinja dobro jela, nabubri. Ovo stvorenje može da proždire žrtve, što ga premašuje po veličini. Usta mu se mogu vrlo široko otvoriti, ponekad se iz njega jasno vidi dio tijela plijena. Nakon ovakvog spektakla, nema sumnje da je slatkovodna hidra grabežljivac u svom načinu hranjenja.
Način reprodukcije
Ako stvorenje ima dovoljno hrane, razmnožavanje se odvija vrlo brzo pupanjem. Za nekoliko dana mali pupoljak izraste u potpuno formiranu jedinku. Često se na hidrinom tijelu pojavljuje nekoliko takvih pupoljaka, koji se potom odvajaju od majčinog tijela. Ovaj proces se naziva aseksualna reprodukcija.
U jesen, kada voda postane hladnija, slatkovodna stvorenja mogu se razmnožavati seksualno. Ovaj proces funkcionira na sljedeći način:
- Gonade se pojavljuju na tijelu pojedinca. Neki od njih proizvode muške stanice, dok drugi proizvode jajašca.
- Muške reproduktivne ćelije kreću se u vodi i ulaze u tjelesnu šupljinu hidri, oplođujući jajašca.
- Kada se formiraju jaja, hidra najčešće umire, a iz jaja se rađaju nove jedinke.
 U prosjeku, dužina tijela hidre je od 1 mm do 2 cm, ali može biti i malo više.
U prosjeku, dužina tijela hidre je od 1 mm do 2 cm, ali može biti i malo više. Nervni sistem i disanje
U jednom od slojeva tijela ovog stvorenja nalazi se raštrkani nervni sistem, au drugom mali broj nervnih ćelija. Ukupno u tijelu životinje postoji 5 hiljada neurona. Životinja ima nervne pleksuse blizu usta, na tabanu i na pipcima.
Hidra ne dijeli neurone u grupe. Ćelije percipiraju iritaciju i šalju signal mišićima. Nervni sistem pojedinca sadrži električne i hemijske sinapse, kao i opsin proteine. Govoreći o tome šta hidra diše, vrijedi spomenuti da se proces izlučivanja i disanja odvija na površini cijelog tijela.
Regeneracija i rast
Ćelije slatkovodni polip su u procesu stalnog ažuriranja. U sredini tijela se dijele, a zatim prelaze na pipke i taban, gdje umiru. Ako ima previše ćelija koje se dijele, one se kreću u donji dio tijela.
Ova životinja ima neverovatnu sposobnost regeneracije. Ako mu presiječete torzo poprečno, svaki dio će biti vraćen na prethodni izgled.
 Ćelije slatkovodnog polipa su u procesu stalnog obnavljanja.
Ćelije slatkovodnog polipa su u procesu stalnog obnavljanja. Životni vijek
U 19. veku se mnogo pričalo o besmrtnosti životinja. Neki istraživači su pokušavali da dokažu ovu hipotezu, dok su drugi želeli da je opovrgnu. Godine 1917., nakon četverogodišnjeg eksperimenta, teoriju je dokazao D. Martinez, zbog čega je hidra službeno postala vječno živo biće.
Besmrtnost je povezana sa neverovatna sposobnost do regeneracije. Smrt životinja u zimsko vrijeme povezano sa nepovoljni faktori i nedostatak hrane.
Slatkovodne hidre su zabavna bića. Četiri vrste ovih životinja nalaze se širom Rusije i svi su slični jedni drugima. Najčešće su obične i stabljikaste hidre. Kada se kupate u rijeci, na njenoj obali možete pronaći cijeli tepih ovih zelenih stvorenja.