В связи с выходом на сушу и более активной жизнедеятельностью характерной для высших позвоночных, все отделы мозга пресмыкающихся достигают более прогрессивного развития.
1. Передний мозг значительно преобладает над другими отделами. Мантия остается тонкой, но на ее поверхности местами появляются медиальное и латеральное скопления нервных клеток – серое вещество, представляющее зачаточную кору больших полушарий. У рептилии кора еще не играет роли высшего отдела мозга, она является высшим обонятельным центром. Но в процессе филогенеза, разрастаясь и принимая другие виды чувствительности, помимо обонятельной, она привела к возникновению коры головного мозга млекопитающих. Полушария переднего мозга пресмыкающихся полностью прикрывают промежуточный мозг. Роль высшего интегративного центра выполняют полосатые тела (зауропсидный тип мозга)
2. Промежуточный мозг образован зрительными буграми и подбугровой областью. На дорсальной его стороне находится эпифиз и особый теменной орган, имеющий глазоподобное строение у ящериц. На вентральной стороне находится гипофиз.
3. Средний мозг довольно большой, имеет вид двухолмия. Это центр зрительных восприятий, приобретающий большое значение для наземных животных.
4. Мозжечок имеет вид полукруглой пластинки, развит слабо, но лучше, чем у амфибий, в связи с усложнением координации движений.
5. Продолговатый мозг образует резкий изгиб, характерный для высших позвоночных. От его ядер берут начало черепно-мозговых нервов.
Всего у рептилий 12 пар черепно-мозговых нервов.
ГОЛОВНОЙ МОЗГ ПТИЦ

Эволюция головного мозга позвоночных: а - рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее; 1 - обонятельные доли; 2 - передний мозг; 3 - средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - промежуточный мозг
1.Передний мозг развит хорошо, полушария имеют значительную величину, частично прикрывают промежуточный мозг. Но увеличение полушарий происходит за счет развития полосатых тел (зауропсидный тип мозга), а не коры. Обонятельные доли очень малы, так как обоняние теряет ведущее значение.
2 Промежуточный мозг мал, прикрыт полушариями переднего мозга. На дорсальной стороне его находится эпифиз (развит слабо), а на вентральной – гипофиз.
3. Средний мозг довольно большой, за счет крупных зрительных долей (двухолмие), что связано с прогрессивным развитием зрения.
4. Мозжечок сильно развит в связи со сложной координацией движений при полете. Он имеет поперечную исчерченность, и свою кору.
5. Продолговатый мозг содержит скопление нервных клеток в виде ядер, от которых берут начало черепно-мозговые нервы от 5-й до 12-й пары.
Всего 12 пар черепно-мозговых нервов.
ГОЛОВНОЙ МОЗГ МЛЕКОПИТАЮЩИХ

Эволюция головного мозга позвоночных: а - рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее; 1 - обонятельные доли; 2 - передний мозг; 3 - средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - промежуточный мозг
1 Передний мозг достигает особенно больших размеров, прикрывая остальные отделы мозга. Его увеличение происходит за счет коры, которая становится главным центром высшей нервной деятельности (маммальный тип мозга). Площадь коры увеличивается за счет образования извилин и борозд. Спереди от больших полушарий у большинства млекопитающих (кроме китообразных, приматов и, в том числе, человека) расположены крупные обонятельные доли, что связано с большим значением обоняния в жизни зверей.
2 Промежуточный мозг, образованный зрительными буграми (thalamus) и подбугровой областью (hypothalamus), скрыт полушариями переднего мозга. На дорсальной его стороне находится эпифиз, а на вентральной – гипофиз.
3 Средний мозг прикрыт полушариями переднего мозга, отличается сравнительно небольшими размерами и представлен не двухолмием, а четверохолмием. Полость среднего мозга, или сильвиев водопровод, представляет собой лишь узкую щель.
4 Мозжечок сильно развит и имеет более сложное строение; состоит из центральной части – червя с поперечными бороздами и парных полушарии. Развитие мозжечка обеспечивает сложные формы координации движений.
5 Продолговатый мозг частично прикрыт мозжечком. Отличается от представителей других классов тем, что потоком четвертого желудочка обособляются продольные пучки нервных волокон – задние ножки мозжечка, а на нижней поверхности имеются продольные валики – пирамиды. От головного мозга отходит 12 пар черепно-мозговых нервов
48. 50. Филогенетически сложившиеся типы и формы иммунного ответа. Характеристика особенностей иммунной системы позвоночных.
Филогенез иммунной системы.
Иммунная система осуществляет защиту организма от проникновения в организм генетически чужеродных тел: микроорганизмов, вирусов, чужих клеток, инородных тел. Ее действие основано на способности отличать собственные структуры от генетически чужеродных, уничтожая их.
В эволюции сформировалось три главных формы иммунного ответа:
1) 1.Фагоцитоз, или неспецифическое уничтожение чужеродного материала;
2) 2.Клеточный иммунитет, основанный на специфическом распознавании и уничтожении такого материала Т-лимфоцитами;
3) 3.Гуморальный иммунитет, осуществляемый путем образования потомками В-лимфоцитов, так называемыми, плазматическими клетками иммуноглобулинов и связывания ими чужеродных антигенов.
В эволюции выделяют три этапа формирования иммунного ответа:
1. 1. Квазииммунное (лат наподобие) распознавание организмов своих и чужеродных клеток. Этот тип реакции наблюдается от кишечнополостных до млекопитающих. Эта реакция не связана с выработкой иммунных тел, и при этом не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала.
2. 2. Примитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами – клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память.
3. 3. Система интегрального клеточного и гуморального иммунитета . Для нее характерны специфические клеточные и гуморальные реакции на чужеродные тела, наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных.
Круглоротые способны формировать антитела, но вопрос о наличии у них вилочковой железы, как центрального органа иммуногенеза, является пока открытым. Впервые тимус обнаруживается у рыб.
Эволюционные предшественники лимфоидных органов млекопитающих – тимус, селезенка, скопление лимфоидной ткани обнаруживаются в полном объеме у амфибий. У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что характерно для птиц и млекопитающих.
Особенность иммунного ответа птиц состоит в налиции особоги лимфоидного органа – фабрициевой сумки. В этом органе образуются В-лимфоциты, которые после антигенной стимуляции способны трансформироваться в плазматические клетки и вырабатывать антитела.
У млекопитающих органы иммунной системы разделяют на два типа: центральные и периферические. В центральных органах созревание лимфоцитов происходит без существенного влияния антигенов. Развитие периферических органов, наоборот, непосредственно зависит от антигенного воздействия – лишь при контакте с антигеном в них начинаются процессы размножения и дифференциации лимфоцитов.
Центральными органами иммуногенеза у млекопитающих являются тимус, где происходит образование и размножение Т-лимфоцитов, а также красный костный мозг, где образуются и размножаются В-лимфоциты.
На ранних стадиях эмбриогенеза и желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфотические клетки. После рождения источником стволовых клеток становится красный костный мозг.
Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще практически не сформированы и образование в них лимфоцитов начинается только после антигенной стимуляции, после того, как они заселяются Т- и В-лимфоцитами из центральгых органов иммуногенеза.
49. 51. Онтогенез, его типы и периодизация.
Онтогенез, или индивидуальное развитие, – это совокупность преобразований, происходящих в организме от момента образования зиготы до смерти. Термин «онтогенез» впервые введен биологом Э.Геккелем в 1866 г. (от греч. онтос- существо и генезис- развитие).
Учение об онтогенезе – это один из разделов биологии, который изучает механизмы, регуляцию и особенности индивидуального развития организмов.
Знание онтогенеза имеет не только общетеоретическое значение. Оно необходимо врачам для понимания особенностей течения патологических процессов в разные возрастные периоды, профилактики заболеваний, а также для решения социально- гигиенических проблем, связанных с организацией труда и отдыха людей различных возрастных групп.
Различают 2 типа онтогенеза: непрямой и прямой. Непрямой протекает в личиночной форме. Личинки ведут активный образ жизни, сами себе добывают пропитание. Для осуществления жизненных функций у личинок имеется ряд провизорных (временных) органов, отсутствующих у взрослых организмов. Этот тип развития сопровождается метаморфозом (превращением) -анатомо-физиологической перестройкой организма. Он свойствен различным группам беспозвоночных (губкам, кишечнополостным, червям, насекомым) и нисшим позвоночным (амфибиям).
Прямое развитие может протекать в неличиночной форме или быть внутриутробным. Неличиночный тип развития имеет место у рыб, пресмыкающихся, птиц, а также беспозвоночных, яйцеклетки которых богаты желтком - питательным материалом, достаточным для завершения онтогенеза. Для питания, дыхания и выделения у зародышей также развиваются провизорные органы.
Внутриутробный тип развития характерен для млекопитающих и человека. Их яйцеклетки почти не содержат питательного материала, и все жизненные функции осуществляются через материнский организм. В связи с этим у зародышей имеются провизорные органы – зародышевые оболочки и плацента, обеспечивающая связь организма матери и плода. Это наиболее поздний в филогенезе тип онтогенеза, и он обеспечивает наилучшим образом выживание зародышей.
Онтогенез включает в себя ряд преемственно связанных и в основных чертах генетически запрограммированных периодов:
1. Предэмбриональный (он же проэмбриональный, или предзиготный период, или прогенез);
2. Эмбриональный (или антенатальный для человека) период;
3. Постэмбриональный (или постнатальный для человека) период.
a. 52. Общая характеристика предзиготного периода, стадии эмбрионального развития. Критические периоды. Тератогенные факторы.
ПРЕДЗИГОТНЫЙ ПЕРИОД
Этот период протекает в организме родителей и выражается в гаметогенезе – образовании зрелых яйцеклеток и сперматозоидов.
В настоящее время известно, что в этот период происходит ряд процессов, имеющих прямое отношение к ранним стадиям эмбрионального развития. Так, в ходе созревания яйцеклеток в пахинеме мейоза наблюдается амплификация генов (образование многочисленных копий), отвечающих за синтез р-РНК, с последующим выделением их из ДНК и накоплением вокруг ядрышек. Эти гены включаются в транскрипцию на ранних стадиях эмбриогенеза, обеспечивая накопление р-РНК, участвующей в образовании рибосом. Кроме того, в предзиготном периоде происходит также накопление как бы впрок и-РНК, включающейся в биосинтез белка только на ранних стадиях дробления зиготы.
Во время овогенеза в яйцеклетках идет накопление желтка, гликогена и жиров, которые расходуются в процессе эмбриогенеза.
По количеству содержания желтка (lecithos) яйцеклетки могут быть:
· олиголецитальными (маложелтковыми);
· мезолецитальными (со средним количеством желтка);
· полилецитальными (многожелтковые).
По характеру распределения желтка в цитоплазме яйцеклетки бывают:
· изолецитальные (греч. Isos – равный, желток распределен в клетке равномерно);
· телолецитальные (греч. thelos – конец, желток смещен ближе к вегетативному полюсу, а клеточное ядро – к анимальному);
· центролецитальные (желток располагается в центральной части яйцеклетки)
Изолецитальные клетки характерны для ланцетника и млекопитающих, телолецитальные – для амфибий (умеренно телолецитальные, для рептилий и птиц – резкотелолецитальные), центролецитальные – для насекомых.
Яйцеклетки некоторых видов животных еще до оплодотворения приобретают билатеральную симметрию, однако она еще неустойчива и может в дальнейшем переориентироваться.
У многих видов животных еще до оплодотворения начинается сегрегация (перераспределение) органоидов и включений в яйцеклетках; отмечается скопление гликогена и и РНК на анимальном полюсе, комплекса Гольджи и аскорбиновой кислоты - на экваторе. Сегрегация продолжается и после оплодотворения.
ЭМБРИОНАЛЬНЫЙ ПЕРИОД
Эмбриональный период начинается с зиготы и заканчивается либо выходом молодых особей из яйцевых оболочек, либо рождением нового организма. Этот период состоит из стадий: зиготы, дробления, гаструляции и гисто- и органогенеза.
ХАРАКТЕРИСТИКА СТАДИЙ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
НА ПРИМЕРЕ ЧЕЛОВЕКА.
После оплодотворения наступает первая стадия эмбрионального развития - стадия зиготы (стадия одноклеточного зародыша). Зигота, будучи одной клеткой, имеет потенции к развитию целостного многоклеточного организма, т.е. обладает тотипотентностью.
Стадия дробления: начиная с этой стадии, зародыш становится многоклеточным, но по размерам практически не превышает зиготу. Дробление заключается в том, что хотя клетки делятся митозом, они не вырастают до размеров материнских клеток, т.к. у них отсутствует гетеросинтетическая интерфаза, а период G1 аутосинтетической интерфазы приходится на телофазу предшествующего деления. Стадия дробления заканчивается образованием бластулы. Первые бластомеры, как и зигота, обладают свойством тотипотентности, что служит основой рождения монозиготных (однояйцевых) близнецов.
У человека бластула образуется на 6-7 день развития и имеет вид пузырька (бластоциста), стенки которого образованы одним слоем клеток - трофобластом, выполняющих функции питания и выделения. Внутри пузырька имеется скопление клеток - эмбриобласт, из которого в дальнейшем развивается тело зародыша.
На стадия гаструляции (у человека с 7 по 19 день) происходит образование зародышевых листков (эктодермы, энтодермы и мезодермы), и закладывается комплекс осевых органов (хорда, нервная трубка и кишечная трубка).
В период гистогенеза и органогенеза идет закладка временных (провизорных) и окончательных (дефинитивных) органов. У позвоночных животных, в том числе и у человека, провизорные органы называются зародышевыми оболочками. Для всех позвоночных характерно развитие желточного мешка. У рыб, амфибий, рептилий и птиц он содержит желток и выполняет трофическую и кроветворную функции. У истинно наземных животных, кроме желточного мешка, имеется также амнион, наполненный жидкостью, создающей водную среду для развития зародыша. Позвоночные, имеющие амнион (рептилии, птицы и млекопитающие), называются амниотами , а не имеющие его - анамниями (рыбы, амфибии).
У рептилий и птиц, кроме желточного мешка и амниона, закладываются: аллантоис (мочевой мешок, накапливающий мочевину) и серозная оболочка (обеспечивает дыхание зародыша). У млекопитающих вместо серозной оболочки образуется хорион (ворсинчатая оболочка), который обеспечивает зародышу питание, дыхание и выделение. Хорион образуется из трофобласта и соединительной ткани. Со стадии плацентации он участвует в образовании плаценты. Амнион содержит околоплодные воды. В желточном мешке образуются первые кровеносные сосуды и первые клетки крови. Аллантоис у млекопитающих и человека определяет место расположения плаценты.
Гисто- и органогенез у человека начинается на четвертой неделе и заканчивается к рождению.
Вначале из так называемой первичной эктодермы вычленяются клетки, образуя нервную пластинку, из которой в дальнейшем развиваются все органы нервной системы и часть органов чувств. Из оставшейся вторичной эктодермы закладываются эпидермис и его производные - сальные, потовые, молочные железы, ногти, волосы и некоторые другие образования.
Из энтодермы формируются: эпителий желудочно-кишечного тракта, дыхательных путей, печень и поджелудочная железа.
Из мезодермы - скелет, поперечнополосатая и гладкая мускулатура, сердечно-сосудистая система и основная часть мочеполовой системы.
КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
В 1921 г. Стоккард Ц.Р. положил начало представлениям о так называемых критических периодах развития животных организмов. Этой проблемой позже у нас в стране занимался Светлов П.Г., который в 1960 г. сформулировал теорию критических периодов развития, проверил ее экспериментально. Сущность ее состоит в том, что каждый этап развития зародыша начинается коротким периодом качественно новой перестройки, сопровождающемся детерминацией, пролиферацией и дифференцировкой клеток. В этот период наблюдается особая восприимчивость к различным повреждающим факторам среды – физическим, химическим и в ряде случаев – биологическим, которые могут ускорять, замедлять и даже приостанавливать развитие.
В онтогенезе человека выделяют следующие критические периоды: 1) гаметогенез; 2)оплодотворение; 3)имплантацию; 4)развитие комплекса осевых органов и формирование плаценты (3-8-я недели) 5) периоды дифференцировки того или иного органа или системы органов, (20-24-я недели);. 6)рождение; 7) период новорожденности (до 1 года); 8) половое созревание.


b. 53. Основные механизмы эмбриогенеза.
ОБЩИЕ МЕХАНИЗМЫ ЭМБРИОГЕНЕЗА
1.Деление клеток
2. Клеточная дифференцировка
3. Дифференциальная активность генов
4. Эмбриональная индукция
5. Межклеточные взаимодействия
6 Миграция клеток.
7.Гибель клеток
8. Клональный принцип развития
9. Рост.
10. Морфогенез.
В основе эмбрионального развития лежат разнообразные процессы /механизмы/, к которым относятся: клеточные деления, дифференцировка, эмбриональная индукция, межклеточные взаимодействия, миграция клеток, гибель клеток, клональный принцип развития, рост, морфогенез и дифференциальная активность генов.
1.Деление клеток лежит в основе пролиферации /разрастания клеток/ и является основным механизмом обеспечения роста, то есть увеличения массы и размеров тела. Кроме того, в ходе клеточных делений в ряде случаев происходит переключение генетических программ и, как следствие этого, - специализация клеток для выполнения определенных функций.
2. Клеточная дифференцировка - это процесс, когда из внешне однообразных клеток и их комплексов возникают специализированные клетки, отличающиеся от материнских морфологическими и функциональными особенностями. Этот процесс носит дивергентный /разнонаправленный/ характер. С биохимической точки зрения, дифференцировка - это выбор из некоего множества возможных путей биосинтеза какого-либо одного (например, клетками-предшественниками эритроцитов выбор пути синтеза гемоглобина, а клетками хрусталика глаза - белка кристалина). С морфологической точки зрения, дифференцировка выражается в приобретении специфических черт строения.
В результате дифференцировки развивается популяция высокоспециализированных клеток либо утративших ядра /эритроциты, ороговевшие клетки эпидермиса/, либо в клетках начинается синтез высокоспецифичных веществ, например, сократимых белков актина и миозина – в мышечных волокнах, определенных гормонов – в клетках желез внутренней секреции, и т. д.
Путь, по которому должна идти дифференцировка тех или иных клеток, генетически детерминирован /предопределен/. На стадии дробления детерминация клеток носит еще неустойчивый характер /лабильна/, и направление дифференцировки можно изменить. Это подтвердил в первой четверти ХХ века Ганс Шпеман в экспериментах на тритонах. Он пересаживал эктодермальные клетки, взятые у тритона одного вида, в энтодерму другого. И хотя клетки донора отличались по цвету от клеток реципиента, они развивались в те же зачатки, что и окружавшие их клетки реципиента. Если донорами были организмы, закончившие процесс гаструляции, то эктодермальные клетки нервной пластинки, пересаженные в кожу, давали зачаток нервной ткани, то есть путь их дифференцировки был уже предопределен.
Сейчас известны некоторые факторы, определяющие дифференцировку тканей. Самым ранним фактором, который проявляется уже на стадии бластулы, является сегрегация /лат. «отделение»/ цитоплазматических структур зиготы, в силу чего во время дробления в первые бластомеры попадают отличающиеся друг от друга участки цитоплазмы. Таким образом, видимо, незначительные качественные различия, имеющиеся в разных участках цитоплазмы ооцитов, влияют на судьбу бластомеров. Есть также данные, что дифференцировка многих тканей зародыша может происходить лишь при наличии некоторого критического числа клеток.
Основным механизмом дифференцировки клеток является дифференциальная активность генов.
3. Эмбриональная индукция – это влияние одной ткани или зачатка органа зародыша /индуктора/ на закладку других зачатков органов. Так, например, у позвоночных закладка хордо-мезодермального комплекса индуцирует /побуждает к развитию/ закладку нервной трубки.
Другой формой индуцирующих воздействий являются межклеточные взаимодействия.
4. Межклеточные взаимодействия осуществляются посредством щелевых контактов, где плазматическая мембрана одних клеток вступает в тесный контакт с плазмалеммой других клеток. В области этих контактов между клетками может передаваться слабый электрический ток, ионы неорганических веществ или даже относительно крупные молекулы органических веществ.
5 Миграция клеток. В ходе эмбриогенеза происходит миграция как отдельных клеток, так и их комплексов, на различные расстояния. Отдельные клетки обычно мигрируют при помощи амебоидного движения, обследуя при этом непрерывно свое окружение.
6.Гибель клеток (апоптоз) является необходимым процессом многих стадий развития зародыша. Так, разделению пальцев ног и рук предшествует гибель клеток, расположенных в межпальцевых промежутках.
7. Клональный принцип развития. Экспериментальным путем показано, что многим клеткам раннего зародыша не суждено участвовать в дальнейшем развитии. Многие структуры зародыша строятся из клеток, которые развиваются в ходе деление только отдельного, небольшого числа клеток.
8. Рост. Под ростом понимается увеличение массы тела и его размеров. Рост носит неравномерный характер, разные ткани и разные части зародыша растут с разной скоростью.
9. Морфогенез. Это процесс пространственного становления внешней и внутренней конфигурации частей тела и органов зародыша. Общепринятой теории, объясняющей механизмы этого процесса, пока нет. Наиболее подходящей является концепция позиционной информации , предложенная Л. Вольпертом /1975/,согласно которой клетки способны воспринимать позиционную информацию, которая содержит указание о местоположении клеток относительно других клеток и тем самым определяется план, в соответствии с которым происходит развитие зародыша.
,
рептилии (пресмыкающиеся) ,
птицы, их гнезда, яйца и голоса , и
млекопитающие (звери) и следы их жизнедеятельности ,
20
цветных ламинированных определительных таблиц
, в том числе:
водные беспозвоночные ,
дневные бабочки ,
рыбы ,
амфибии и рептилии ,
зимующие птицы ,
перелетные птицы ,
млекопитающие и их следы ,
4
карманных полевых определителя
,
в том числе:
обитатели водоемов ,
птицы средней полосы и
звери и их следы , а также
65
методических пособий
и
40
учебно-методических фильмов
по методикам
проведения научно-исследовательских работ в природе (в полевых условиях).
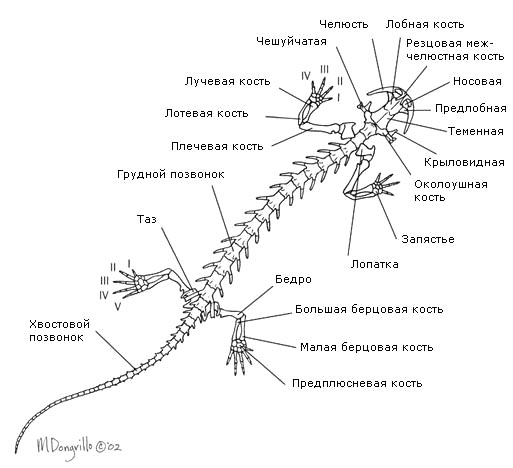
Анатомия амфибий: общий обзор
Строение тела или анатомия.
Тело
разделено на голову, туловище, хвост
(только у хвостатых и безногих) и конечности,
которые могут и отсутствовать. Голова
подвижна, соединена с туловищем. Скелет и
позвоночник разделён на отделы
. К туловищным
позвонкам прикрепляются рёбра, если они есть.
Земноводные имеют две пары первичнопятипалых конечностей ; зачаточные формы шейной части обеспечивают им возможность самостоятельного движения головой.
Кожа голая, лишенная чешуи. Эпидермис богат многоклеточными железами, которые обеспечивают наличие на поверхности кожи жидкостной пленки, без которой при кожном дыхании невозможен газообмен. Эпидермис многослойный, кориум тонкий, но обильно насыщенный капиллярами.
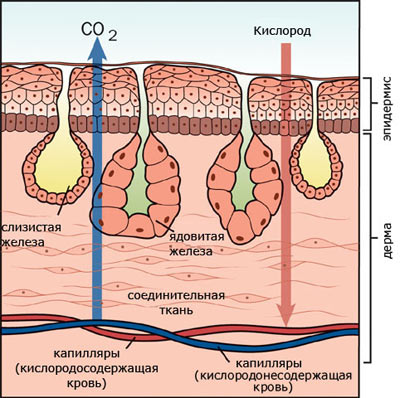
В нижних слоях эпидермиса и в кориуме расположены пигментные клетки , обусловливающие видоспецифичность окраски.
Скелет конечностей образован скелетом пояса конечностей и скелетом свободных конечностей.
Плечевой пояс лежит в толще мускулатуры и включает парные лопатки, ключицы и вороньи кости, соединённые с грудиной. Скелет передней конечности состоит из плеча (плечевая кость), предплечья (лучевая и локтевая кости) и кисти (кости запятстья, пястья и фаланги пальцев).
Тазовый пояс состоит из парных подвздошных седалишных и лобковых костей, сросшихся между собой. Он прикреплен к крестцовому позвонку через подвздошные кости. В состав скелета задней конечности входят бедро, голень (большая и малая берцовая кости) и стопа. Кости предплюсны, плюсны и фаланги пальцев. Крестец состоит из одного только позвонка.
Двигательная система.
Характер движения земноводных довольно однообразен
и может быть сведен к двум основным типам.
Ископаемые и современные хвостатые земноводные сохранили свойственный рыбам основной тип движения - при помощи сильных боковых изгибов всего тела, но с опорой на короткие ножки при движении по земле. При коротких конечностях боковые изгибы туловища увеличивают длину шага, а изгибы хвоста помогают сохранять равновесие. При движении в воде конечности сколько-нибудь заметной роли не играют. С помощью изгибов всего тела передвигаются и безногие.
Бесхвостые земноводные по суше передвигаются прыжками , поднимая тело в воздух резким толчком обоих задних конечностей. Коротконогие виды, например жабы , помимо прыжков могут медленно шагать , последовательно переставляя конечности.
В воде бесхвостые плавают , энергично работая задними конечностями (стиль "брасс", но без участия передних конечностей). Предполагают, что мощные задние конечности развились в результате приспособления к плаванию, и лишь позднее использованы для прыжков на суше.
У амфибий довольно крупная, широкая голова , которая переходит непосредственно в широкое и короткое тело . Лобные и теменные кости слиты в парную лобнотеменную кость. В черепе характерно то, что челюстно-нёбный аппарат и квадратная кость соединены с черепом неподвижно; два мыщелка черепа принадлежат собственно слившемуся с ним первому шейному позвонку, так что первый позвонок амфибий есть в сущности второй.
Мозг
амфибий отличается от мозга рыб
большим развитием переднего отдела (передний
мозг
), содержащего большое число нервных
клеток (серое вещество). Полушария
переднего
мозга малы и полностью разделены. Отделы мозга
лежат в одной горизонтальной плоскости. Обонятельные
доли сильно развиты. Мозжечок
развит очень
слабо в связи с малой подвижностью и
однообразным характером движений.
Черепномозговых нервов 10 пар. У личинок имеются
органы боковой линии
.
Спинной мозг
развит лучше головного.
Головной мозг состоит из 5 отделов
: передний
мозг, промежуточный, продолговатый, средний,
мозжечок. Промежуточный
мозг хорошо развит. Продолговатый
мозг является центром дыхательной, кровеносной и
пищеварительной системы. Средний
мозг
относительно невелик.
Органы осязания хорошо развиты. Органы боковой линии сигнализируют амфибиям о волнообразных колебаниях воды. Они даны им для активной локации водного пространства, особенно в мутной воде или в ночное время, и полностью заменяют зрение. Являясь органами дистанционного осязания, такие живые приборы чувствуют и колебания, вызванные движениями подводных жителей. Органы боковых линий находятся на поверхности кожи земноводных, живущих исключительно в воде, и для каждого вида имеются свои особенности.
Органом осязания служит вся кожа , в которой имеются осязательные нервные окончания.
Во рту также находятся органы осязания в виде вкусовых луковичек . Зубы могут как присутствовать у некоторых видов, так и отсутствовать. Зубы, как и у пресмыкающихся, приспособлены лишь к схватыванию и к удерживанию добычи, но не могут служить для ее разжевывания. Звуки могут издавать только бесхвостые амфибии , да и то преимущественно самцы.
Носовая полость снабжена задними носовыми отверстиями и носослезными каналами.
Глаза похожи на глаза рыб, но не имеют серебристой оболочки, ни отражательной, ни серповидного отростка. Аккомодация глаза осуществляется перемещением хрусталика. Глаза приспособлены к видению на большом расстоянии. Слезные железы отсутствуют, но есть Гардерова железа, секрет которой смачивает роговицу и предохраняет её от высыхания. Роговица выпуклая. Хрусталик имеет форму двояковыпуклой линзы, диаметр которой меняться в зависимости от освещения; аккомодация происходит засчёт изменения расстояния хрусталика до сетчатки. У многих развито цветное зрение .

Строение уха различается у бесхвостых и хвостатых амфибий.
Мускулатура подразделяется на мускулатуру туловища и конечностей. Туловищная мускулатура сегментирована. Группы специальных мышц обеспечивают сложные движения рычажных конечностей. На голове расположены мышцы поднимающие и опускающие. Благодаря сокращениям мышц или групп мышц амфибии могут совершать сложные движения. Особенно хорошо развиты мышцы конечностей.
Пищеварительная система земноводных имеет почти такое же строение, как у рыб. Все земноводные питаются только подвижной добычей . На дне ротоглоточной полости находятся язык. В ротоглоточную полость открываются протоки слюнных желез, секрет которых не содержит пищеварительных ферментов. Из ротоглоточной полости пища по пищеводу поступает в желудок, оттуда в двенадцатипёрстную кишку. Сюда открываются протоки печени и поджелудочной железы. Переваривание пищи происходит в желудке и в двенадцатипёрстной кишке. Тонкий кишечник переходит в толстый, заканчивающийся прямой кишкой, которая образует расширение - клоаку. В отличие от рыб задняя кишка открывается не прямо наружу, а в особое ее расширение, называемое клоакой. В клоаку открываются также мочеточники и выводные протоки органов размножения.

Органами дыхания у земноводных являются:
- лёгкие (специальные органы воздушного дыхания);
- кожа и слизистая выстилка ротоглоточной полости (дополнительные органы дыхания);
- жабры (у некоторых водных обитателей и у головастиков).
У большинства видов (кроме безлёгочных саламандр) имеются лёгкие небольшого объёма, в виде тонкостенных мешков, оплетённых густой сетью кровеносных сосудов. Каждое лёгкое открывается самостоятельным отверстием в гортанно-трахейную впадину (здесь расположены голосовые связки, открывающиеся щелью в ротоглоточную полость). Воздух нагнетается в лёгкие за счёт изменения объема ротоглоточной полости: воздух поступает в ротоглоточную полость через ноздри при опускании её дна. При поднимании дна воздух проталкивается в лёгкие.
Горло несколько раз в секунду оттягивается вниз , благодаря чему в ротовой полости создается разреженное пространство. Тогда воздух проникает через ноздри в ротовую полость, а оттуда в легкие. Обратно он выталкивается под действием мускулатуры стенок тела. Погружённая в воду амфибия целиком переходит на кожное дыхание .
Кровеносная система замкнутая, состоит из большого и малого кругов кровообращения. Появление второго круга связано с приобретением лёгочного дыхания. В организме есть кожнолёгочные артерии (несут венозную кровь к лёгким и коже), сонные артерии (снабжены артериальной кровью органы головы), дуги аорты несут смешанную кровь к остальным органам тела.

I - венозная пазуха; II - правое предсердие; III - левое предсердие; IV - желудочек; V - артериальный ствол;
1 - лёгочно-кожная артерия; 2 - дуга аорты; 3 - сонная артерия; 4 - язычная артерия; 5 - сонная железа; 6 - подключичная артерия; 7 - общая аорта; 8 - кишечная артерия; 9 - кожная артерия; 10 - лёгочная вена; 11 - лёгкое; 12 - задняя полая вена; 13 - кожная вена; 14 - брюшная вена; 15 - печень; 16 - почечная вена.
Малый круг кровообращения - лёгочный, начинается кожно-лёгочными артериями, несущими кровь к органам дыхания (лёгким и коже); от лёгких обогащённая кислородом кровь собирается в парные лёгочные вены, впадающие в левое предсердие.
Большой круг кровообращения начинается дугами аорты и сонными артериями, которые ветвятся в органах и тканях. Венозная кровь по парным передним полым венам и непарной задней полой вене попадает в правое предсердие. Кроме того, в передние полые вены попадает окисленная кровь, и поэтому кровь в правом предсердии смешанная. Так как органы тела снабжаются смешанной кровью, у амфибий низкий уровень обмена веществ и поэтому они хладнокровные животные.
Аорта проходит в жаберные дуги и разветвляется сначала в наружных жабрах, а позднее и во внутренних. Обратно кровь течет по вене, идущей вдоль хвоста, а затем разветвляется на поверхности желточного пузыря и через желточные вены возвращается обратно в предсердие. Позднее постепенно образуются воротные системы печени и почек. В конце личиночной стадии жаберное дыхание мало-помалу заменяется легочным; передние жаберные дуги превращаются в головные артерии, а средние образуют аорту.
Сердце трёхкамерное. Состоит из двух предсердий (в правом предсердии кровь смешанная, преимущественно венозная, а в левом - артериальная) и одного желудочка. Внутри стенки желудочка образуют складки, препятствующие смешиванию артериальной и венозной крови. Из желудочка выходит артериальный конус, снабжённый спиральным клапаном.
Правое предсердие получает венозную кровь, левое - артериальную (из лёгких и кожи). Венозная и артериальная кровь лишь частично смешиваются в полости желудочка, стенки которого имеют сложную систему мускульных перекладин. В лёгочные вены направляется главным образом венозная кровь, дуги аорты заполняются смешанной кровью, и лишь сонные артерии получают артериальную кровь.
Сердце образуется у личинок очень рано и тотчас же начинает действовать. Первоначально оно представляет простой мешок, который впоследствии разделяется на отдельные части.
Органы выделения - парные туловищные почки, от которых отходят мочеточники, открывающиеся в клоаку. В стенке клоаки имеется отверстие мочевого пузыря, в который стекает моча, попавшая в клоаку из мочеточников. В туловищных почках не происходит обратного всасывания воды. После наполнения мочевого пузыря и сокращения мышц его стенок, концентрированная моча выводится в клоаку и выбрасывается наружу. Часть продуктов обмена и большое количество влаги выделяется через кожу. Эти особенности не позволили земноводным полностью перейти к наземному образу жизни. У личинок на ранних стадиях развития функционирует т. н. головная почка, или предпочка. Также у всех амфибий лопастная печень, желчный пузырь, поджелудочная железа.
Половая система. Все земноводные раздельнополые. У большинства земноводных оплодотворение наружное (в воде). В период размножения наполненные зрелыми яйцеклетками парные яичники заполняют у самок почти всю брюшную полость. Созревшие икринки выпадают в брюшную полость тела, попадают в воронку яйцевода и, пройдя по нему, через клоаку выводятся наружу. Самцы имеют парные семенники. Отходящие от них семявыводящие канальца попадают в мочеточники, одновременно служащие самцам семяпроводами. Они также открываются в клоаку. Половые клетки по трубчатым протокам попадают в клоаку, а оттуда выбрасываются наружу.
Мышечная система . Отличается от таковой рыбы главным образом большим развитием мускулатуры конечностей и большей дифференцировкой туловищной мускулатуры, состоящей из сложной системы отдельных мышц. В результате первичная сегментация мускулатуры нарушается, хотя в некоторых брюшных и спинных мышцах она еще остается вполне отчетливой.
Нервная система . Головной мозг земноводных отличается от мозга рыб главным образом большим развитием переднего мозга, полным разделением его полушарий и недоразвитым мозжечком, представляющим собой лишь небольшой валик из нервного вещества, прикрывающего переднюю часть IV желудочка. Развитие переднего мозга выражается не только в его увеличении и дифференцировке, но и в том, что, кроме дна боковых желудочков, их бока и крыша содержат нервное вещество, т. е. у земноводных появляется уже настоящий мозговой свод — архипаллиум (из современных рыб архипаллиум имеется у двоякодышащих). Обонятельные доли лишь слабо отграничены от полушарий. Промежуточный мозг сверху лишь слегка прикрыт соседними отделами. К крыше его прикрепляется теменной орган, а от дна отходит хорошо выраженная воронка, к которой прикрепляется гипофиз. Средний мозг хотя и представляет собой значительный отдел, но он относительно меньше, чем у рыб. Недоразвитие мозжечка, как и у двоякодышащих рыб, связано с несложностью движений тела: земноводные— вообще малоподвижные животные, у тех же из них, которые подобно лягушкам могут совершать быстрые движения, они ограничиваются прыжками, т. е. движениями очень простыми. От головного мозга, как и у костистых рыб, отходят только 10 пар головных нервов; XII пара (подъязычный нерв) отходит за пределами черепной коробки, а XI пара (добавочный нерв) вообще не развита.

. I — сверху; II — снизу; III — сбоку; IV — в продольном разрезе (по Паркеру):
1 — полушария переднего мозга, 2 — обонятельная доля, 3 — обонятельный нерв, 4 — промежуточный мозг, 5 — зрительная хиазма, 6 — воронка, 7 — гипофиз, 8 — средний мозг, 9 — мозжечок, 10 — продолговатый мозг, 11 — четвертый желудочек, 12 — спинной мозг, 13 — третий желудочек, 14 — сильвиев водопровод,
III —X — головные нервы, XII — подъязычный нерв

, схема (по Грегори):
1 — черепная коробка, 2 — продолговатый мозг, 3 — слуховой нерв, 4 — полукружные каналы, 5 — полость среднего уха, 6 — евстахиева труба, 7 — глотка, 8 — етремя, 9 — барабанная перепонка
Настоящих спинномозговых нервов у лягушки 10 пар. Три передние пары принимают участие в образовании плечевого сплетения, иннервирующего передние конечности, а четыре задние пары—в образовании пояснично-крестцового сплетения, иннервирующего задние конечности.
Симпатическая нервная система лягушки, как у всех земноводных, развита очень хорошо и представлена главным образом двумя нервными стволами, которые тянутся с обеих сторон позвоночника и образованы цепью нервных узлов, связанных друг с другом тяжами и соединенных со спинномозговыми нервами.
Еще интересные статьи
Тело Земноводные: Подразделяется на голову, туловище и пятипалые конечности. Хвостатые амфибии имеют хвост.
Пресмыкающиеся: Подразделяется на голову, шею, туловище, хвост и пятипалые конечности.
Кожа Земноводные: Тонкая, лишена чешуи, но имеет большое количество желёз, выделяющих слизь.
Пресмыкающиеся: Сухая, лишена желёз и покрыта роговыми чешуями, которые защищают тело от высыхания. Чешуи сдерживают рост, поэтому для рептилий характерна линька.
Позвоночник
Земноводные: 4 отдела: шейной, туловищный, крестцовый и хвостовой. Рёбра редуцированы, у бесхвостых они отсутствуют. Мускулатура не имеет сегментарного строения и представлена дифференцированными группами мышц.
Пресмыкающиеся: 5 отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Имеются рёбра, грудина и грудная клетка. Отделы скелета конечностей те же, что у земноводных. Мускулатура более дифференцирована.
Пищеварительная система Земноводные: Пищеварительная трубка подразделяется на передний, средний и задний отделы. Обособлен желудок. Расширение толстой кишки образует клоаку. Развиты пищеварительные железы.
Пресмыкающиеся: Ротовая полость, глотка, пищевод, желудок, тонкая и толстая кишки. На границе толстой и тонкой кишок находится зачаток слепой кишки. Толстая кишка открывается в клоаку. Развиты пищеварительные железы.
Органы выделения Земноводные: Парные туловищные мочеточники и мочевой пузырь, который открывается в клоаку.
Пресмыкающиеся: Вторичные (тазовые) почки, мочеточники, мочевой пузырь (открывается в клоаку).
Кровеносная система
Земноводные: Сердце трёхкамерное. Два круга кровообращения. По сосудам большого круга течёт смешанная кровь, а головной мозг снабжается артериальной кровью. Земноводные относятся к пойкилотермным животным.
Пресмыкающиеся: Сердце трёхкамерное, но в желудочке имеется неполная перегородка. Два круга кровообращения.
Органы дыхания У взрослых земноводных животных – лёгкие, у личинок – жабры. Дополнительно в дыхании участвует кожа.
Пресмыкающиеся: Лёгкие. Представляют собой растяжимые мешки, внутренняясетка которых имеет сеть перекладин, увеличивающих поверхность. Задний конец трахеи разветвлён на два бронха, которые входят в лёгкие.
Ответить